Last week I introduced the not-so-lovely damaging black fly Simulium damnosum, a vector of the worm that causes river blindness in humans. So what better than introducing this very worm today?
Named Onchocerca volvulus, today’s fellow lacks a common name, but since it is the cause of the disease known as onchocerciasis or river blindness, I think that river-blindness worm is an adequate name.
Adult river-blindness worms live under the skin (in the dermis) of humans, which apparently are the only species that they infect as adults. They have the typical roundworm appearance of any roundworm. Females measure up to 70 cm in length and live inside nodules that they develop under the human skin. Males are much smaller, measuring only 2.3 cm, and can move under the skin from nodule to nodule in search for fertile females.
Three adult males and one adult female of the river-blindness worm. Credits to Hizagi et al. (2014).*
When females are about to release their eggs, they attract males , possibly using pheromones. After mating, the female retains the eggs inside her until the first-stage microfilariae (larvae) hatch. They are then released into the subcutaneous tissue where they wait for a black fly (Simulium damnosum or other species) to bite a human in search for blood. When this happens, the microfilariae end up sucked into the black fly together with the blood.
From the black fly’s gut, the microfilariae migrate to the thoracic flight muscles of the black fly and develop into the first juvenile stage and then into the second. After that they migrate again, now toward the black fly’s proboscis and their salivary glands. In the salivary glands, they mature into the third juvenile stage, which is the infectious stage. The whole cycle inside the blackfly takes about 1 to 3 weeks.
Scan electron microscopy of the head of a female black fly with a juvenile river-blindness worm emerging from her antenna. Credits to the United States Department of Agriculture.
When the black fly bites a human again, the infectious juveniles and delivered via saliva into the bloodstream of the human. From there, they migrate back to the dermis, form nodules and develop into adults over a period of 6 to 12 months, finishing the cycle.
Infected humans develop the disease known as onchocerciasis or river blindness. The main symptom is severe itching, but long-therm infections lead to keratitis (inflammation of the cornea in the eye), which eventually leads to blindness. Onchocerciasis is caused by the microfilariae and the response of the immune system to the infection and not by the adults.
The most effective treatment currently is the use of ivermectin, which kills the microfilariae. It does not kill adults, though, only paralyzing the uterus of females for about 12 to 18 months, preventing them to release more microfilariae during this time. For this reason, a new dose of ivermectin needs to be taken every year as long as adults worms are present. Recent findings, however, suggest that some adult females are developing resistance to ivermectin, which indicates that new treatments need to be developed.
The river-blindness worm is native from tropical Africa, where it probably evolved along with humans. The worms has spread to the Americas, probably during the period of slave trade, and the main non-African countries to have cases of river blindness nowadays are Brazil and Venezuela, but only in isolated regions which are not even close to the dimensions that this disease has in Africa.
River blindness is one of the many neglected tropical diseases, infections that do not receive the adequate investment to allow their control. Despite the considerably good response to the treatment by ivermectin, this is not enough to contain the transmission of the worm, since poor populations that barely have a house to live are even less likely to have the adequate conditions to prevent infections (let alone recurrent infections) in the first place.
Basáñez, M. G., Churcher, T. S., & Grillet, M. E. (2009). Onchocerca–Simulium interactions and the population and evolutionary biology of Onchocerca volvulus. Advances in parasitology, 68, 263-313.
Higazi, T. B., Geary, T. G., & Mackenzie, C. D. (2014). Chemotherapy in the treatment, control, and elimination of human onchocerciasis. Research and Reports in Tropical Medicine, 5, 77-93.
Osei-Atweneboana, M. Y., Awadzi, K., Attah, S. K., Boakye, D. A., Gyapong, J. O., & Prichard, R. K. (2011). Phenotypic evidence of emerging ivermectin resistance in Onchocerca volvulus. PLoS neglected tropical diseases, 5(3), e998.
The evolution of similar traits in distantly related species is common when they have similar lifestyles, and this is particularly noticeable in some parasitic groups. Among flatworms, tapeworms have developed a complex life cycle with two hosts, larval stages and adults that live in the intestine of the definitive hosts where they absorb nutrients directly through the body surface, lacking a digestive system, and have a special structure on their head to remain attached to the host’s guts.
A very similar lifestyle and morphology evolved in a distantly related group of animals, the acanthocephalans or thorny-headed worms. For a long time, the acanthocephalans were considered a phylum of their own, Acanthocephala, but we now know that they are just a group of very specialized rotiferans, whose free-living forms are very small, so small that they are often mistaken for ciliates or other unicelular organisms.
Today’s species is an acanthocephalan that lives very close to humans, so close that they can even live inside us. Named Moniliformis moniliformis, I will call it the “rat acanthocephalan” because, well, it infects rats (and occasionally other mammals including humans).
Adults of the rat acanthocephalan are often found in the intestine of rats but other mammals can also be infected, such as dogs, cats and humans. Males reach up to 5 cm in length while females can be much longer, up to 30 cm.
Adult, probably female, specimen of the rat acanthocephalan.
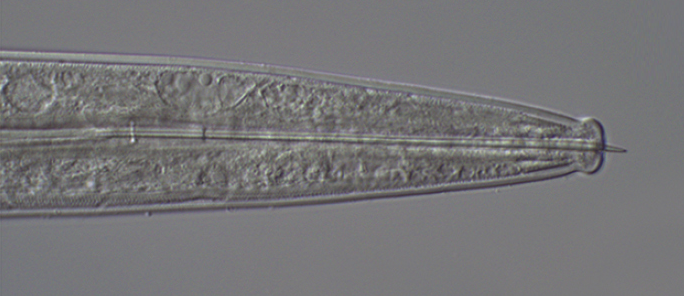
The anterior end of the body has a short cylindrical proboscis covered by hooks, which the animal uses to attach to the host’s intestine. This proboscis is hollow and can be pulled back into the body. There is a septum separating the cavity of the proboscis from the cavity of the rest of the body. Like in tapeworms, the surface of acanthocephalans is covered by a syncytium, a tissue formed by cells that fused together into a single multinucleated structure. Due to the lack of a digestive system, they absorb the nutrients from the hosts intestine directly through their body surface, just like in tapeworms.
Anterior end showing the short proboscis.
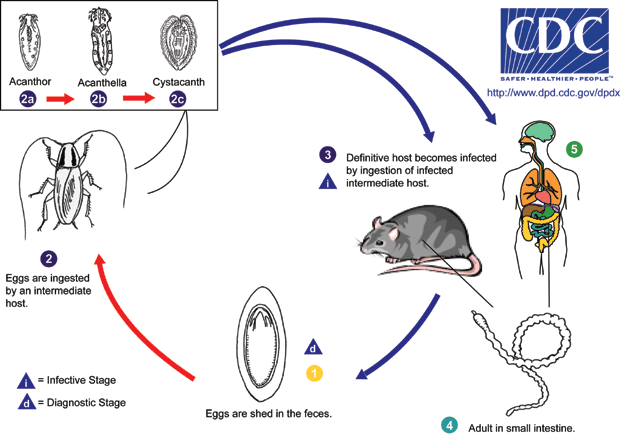
After mating occurs, females release fertilized eggs into the host’s intestine and they leave the body with its feces. The eggs measure about 100 µm in length and 60 µm in width and contain the first larval stage, known as the acanthor. In the environment, the eggs are ingested by the intermediate host, usually a cockroach or sometimes a beetle, and the acanthor hatches, changing into the second-stage larva, the acanthella. After some weeks developing inside the intermediate host, the acanthella changes into the final larval stage, the cystacanth, which forms a cyst inside the intermediate host’s tissues, and there it waits.
An egg under the microscope.
For the cycle to be completed, the intermediate host needs to be eaten by the definitive host. To increase the chances of this happening, the parasite leads to behavioral changes in the intermediate hosts. Infected American cockroaches, for example, show delayed escape responses, increasing the probablity of being captured by a predator. When it happens, the cystacanths are released into the definitive host’s gut and develop into adults.
Life cycle of the rat acanthocephalan.
Humans acting as definitive hosts is a rare occurrence since it requires the ingestion of raw infected cockroaches or beetles. Most reported cases in the literature include small children, which are prone to put everything into their mouths, and the symptoms of the infection include acute abdominal pain and, in very small children, usually less than a year old, more severe symptons such as vomiting, anorexia and diarrhea can also occur. The identification of eggs in stool samples of infected humans is difficult, though, so that the actual infection rate may be much higher than thought, especially in rural areas where insect consumption is a common practice.
Moore J (1983) Altered Behavior in Cockroaches (Periplaneta americana) Infected with an Archiacanthocephalan, Moniliformis moniliformis. Parasitology 69(6):1174–1176. https://doi.org/10.2307/3280893
Salehabadi A, Mowlavi C, Sadjjadi SM (2088) Human Infection with Moniliformis moniliformis (Bremser 1811) (Travassos 1915) in Iran: Another Case Report After Three Decades. Vector-Borne and Zoonotic Diseases 8(1):101–104. http://doi.org/10.1089/vbz.2007.0150
Nematodes are famous because of their parasitic members, which do not only parasitize animals but also plants. People that deal with gardening or agriculture may know that sometimes a plant becomes sick because of “nematodes”.
A genus of nematodes that is commonly associated with grapevines is Xiphinema, whose species are known as dagger nematodes. The two most widely studied species are Xiphinema americanum, the American dagger nematode, and Xiphinema index, the California dagger nematode, but during the last decades it became clear that those species are actually a complex of very similar species and new ones are constantly been described. One of them, described in 2016, is Xiphinema browni, which I decided to call Brown’s dagger nematode. It was named after the nematologist Derek J. F. Brown.
Brown’s dagger nematode was found associated with the roots of grapevines and apple trees in Slovakia and the Czech Republic. Among 86 identified females there was only one male, indicating a huge disparity in sex ratios and the probability that females are parthenogenetic, i.e., they can lay fertile eggs without being fertilized by a male. Females measure up to 2.5 mm in length and the only known male measured 1.8 mm.
Female (left) and male (right) of Xiphinema browni. Modified from Lazarova et al. (2020).*
Since Brown’s dagger nematode was found associated with grapevines, its life cycle is likely similar to that of most other dagger nematodes. Adults are external parasites of grapevine roots and eventually of other woody plants. They live on the root surface and use their long odontostyles (a needle-like proboscis) to perforate the roots and suck the content of their vascular tissue. As a reaction, the plant produces swollen club-like galls on the root tips. The root then branches behind the swollen tip, only to be attacked again, developing another gall and having to branch again. This starts to weaken the plant, which can compromise grape production.
Anterior end of a female with the odontostyle slightly exposed. Modified from Lazarova et al. (2020).*
Females lay their eggs scattered through the soil, not forming clusters, and juveniles pass through about 4 stages in the soil before turning to the parasitic mode.
As another grapevine-feeding dagger nematode, Brown’s dagger nematode is probably also a vector of the grapevine fanleaf virus, which is transmitted to grapevines by the California dagger nematode. This happens when the nematode feeds on an infected plant and then moves to a healthy plant, carrying the virus with it. Grapevine fanleaf causes chlorosis (loss of chlorophyll) and distorts the leaves, making them look like fans, hence the name. As you can imagine, the poor plant becomes even weaker than it already was due to the nematodes sucking it. This can be a nightmare to vineyard owners.
The grapevine fanleaf virus can be a devastating disease for grapevines but in the nematode’s body it seems to have benefitial effects, increasing the survival of these small roundworms. Perhaps this stimulates the dagger nematodes to spread it further, in a sort of “evil coalition”.
Lazarova S, Peneva V, Kumari S (2016) Morphological and molecular characterisation, and phylogenetic position of X.browni sp. n., X.penevi sp. n. and two known species of Xiphinemaamericanum-group (Nematoda, Longidoridae). ZooKeys 574:1–42. https://doi.org/10.3897/zookeys.574.8037
Two mosquitoes of the genus Aedes, Aedes aegypti and Aedes albopictus, are invasive species in tropical and subtropical regions worldwide. While A. aegypti is native from Africa, A. albopictus is originally from southeast Asia, but both species have been spread by humans and continue to increase their range.
Both species are known as vectors of several diseases that affect humans, especially those caused by Flavoviruses, which include the Yellow fever, Dengue fever and Zika fever. Chikungunya, caused by a species of Alphavirus is also transmitted to humans by them. Moreover, they can also transmit some nematodes, such as the heartworm that infects the heart of dogs and other carnivores.
Aedes aegypti biting a human and having a delicious bloody meal. Photo by James Gathany.
Because A. aegypti and A. albopictus pose such a huge threat to public health, getting rid of them is top priority. Here in Brazil, there is a massive national campaign to reduce the ability of Aedes to reproduce by avoiding containers with still water in the open, such as flower vases, buckets, uncovered barrels, discarded tires and virtually everything that can retain water long enough for the larvae to develop. I have to say, though, that this all seems to be useless. The mosquitoes continue to spread and the cases of dengue fever continue to grow. The fact is that the mosquitoes will find a place to lay their eggs. If they don’t find it in your backyard, they will find it in the forest or any vacant lot.
Instead of forcing them to lay their eggs where we cannot see, we should stimulate them to lay their eggs around us and then kill the larvae. Several aquatic predators have been tested as potential allies, including larvivorous fish, dragonfly nymphs, copepods, planarians and even other mosquitoes whoses larvae eat the larvae of Aedes! The use of these predators showed mixed results. Larvivorous fish are difficult to maintain in water tanks at home and dragonfly nymphs are too generalist as predators.
Now a new predator has been suggested: a plant! Yes, a carnivorous plant of the genus Utricularia, which includes species known as bladderworts. These aquatic plants have little bladder-like structures that function as traps to capture small animals. The bladder is hollow and has an internal negative pressure in relation to the environment surrounding it. This negative pressure is created by water being constanly pumped out of the bladder through its walls via active transport. The bladder’s opening is covered by a small lid that avoids water to fill it again when the trap is set. Surrounding the lid, there is a group of bristle-like protuberances. When an animal is moving through the water and moves one of those bristles, they slightly deform the lid, breaking the seal and allowing water to enter the bladder. The negative pressure then sucks water quickly into the bladder, dragging the small animal with it. Then it is only a matter of time for the poor animal to be digested.
Watch the plant in action.
A group of researchers at the University of Rhode Island, USA, tested whether Utricularia macrorhiza, the common North American bladderworth, could be an effective predator of mosquito larvae. By adding U. macrorhiza to containers with larvae of A. aegypti and A. albopictus, they were able to kill 95 to 100% of the larvae in only five days. That’s an amazing result, don’t you think?
Bladderwort with several Aedes larvae (marked with asterisks) in its traps. Credits to Couret et al. (2020).*
Since bladderworts are much easier to raise in tanks and other containers in your backyard than animal predators such as fish and dragonflies, they are a promising new alternative to control the populations of this disease-carrying insects.
So, are you eager to raise some aquatic carnivorous plants to help fight these heinous mosquitoes?
So we are going through a kind of apocalypse as everyone knows. An aggressive and contagious virus has spread all over the world and is causing a major impact in our society, killing thousands of people and crashing the economy.
But I’m not here to talk about how to protect against the virus and who is more vulnerable to it. You can find such information virtually everywhere (but don’t trust the bullshit that Karen the anti-vaxxer or your uncle Donald the boomer is spreading through Whatsapp. That is worse than the virus). Likewise, I will not point out how this pandemic is a direct outcome of our flawed capitalist society and how the fucking rich should be beheaded once and for all. No. I will make a more biological approach and explain a little bit of what this virus is from a structural, functional and taxonomic point of view.
So let’s start with what is a virus.
A virus is basically a parasitic piece of sh… genetic material that infects cells in order to reproduce. Viruses are not quite living beings as they neither have cells nor metabolism. However, they need cells to reproduce. All viruses consists of a strand of nucleic acid (either DNA or RNA) and a capsid, a “box” that protects the nucleic acid. The capsid is usually formed by many copies of one or two proteins that are encoded in the virus’ genetic material. Each individual protein molecule of the capsid is called a capsomere.
Scheme of a helical virus showing the helical capsid in green and the genetic material in blue. Credits to Anderson Brito.*
The Tobacco mosaic virus, that infects tobacco plants and others, has a typical helical capsid.
Most viruses have either a helical or an icosahedral capsid. In a helical capsid, the capsomeres are helically arranged and form an elongate and hollow tube inside of which the genetical material is located. In icosahedral capsids, the capsomeres are arranged to form a icosahedron, i.e., a polyhedron with 20 faces that surrounds the genetic material.
Scheme of an icosahedral virus with an icosahedral capsid (green) surrounding the genetic material (red). Credits to Anderson Brito.*
Adenoviruses are an example of virus with icosahedral capsids. Photo by Graham Beards.**
Many viruses have an additional coat, the envelope, that surrounds the capsid. The envelope is a bi-lipid layer crossed by glycoproteins, like the cell membrane of living organisms, and is formed by the cell membrane or an internal membrane of the cell in which the virus was born. It is, therefore, very similar to the cell membrane of the virus’ host.
Scheme of an enveloped icosahedral virus. The bi-lipid layer is shown in gray and the glycoproteins in orange. Credits to Anderson Brito.*
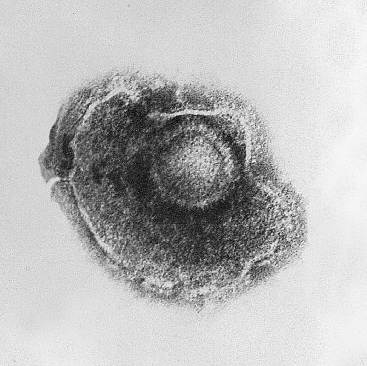
Zika virus (digitally colored blue in this electron microphotograph) is an envoloped icosahedral virus.
Regarding the type of nucleic acid found in viruses, they can be classified into three main groups: DNA viruses, RNA viruses and retroviruses.
DNA viruses have DNA as their nucleic acid. When they infect a cell, they are delivered into the cell’s nucleus, where they depend entirely on the cell’s machinery to reproduce, i.e., they use the hosts DNA-polymerase to produce new DNA strands and the host’s RNA-polymerase to build a viral RNA that will, in turn, be converted into the capsid proteins using the cell’s ribosomes. DNA viruses suffer little mutation because DNA-polymerase enzymes have a proofreading ability, i.e., they can detect errors during replication and fix them. Viruses such as Herpesvirus (which cause herpes and chicken pox), Poxvirus (which include the now extinct Variola virus that caused smallpox) and Adenovirus are all DNA viruses.
The Human alphaherpesvirus 3 (HHV-3) is an enveloped icosahedral DNA virus that causes chickenpox and shingles in humans.
RNA viruses, also called riboviruses, on the other hand, have RNA as their nucleic acid. When they infect a cell, they usually remain in the cell’s cytoplasm. Different from DNA viruses, RNA viruses often have their own RNA-polymerase enzyme and use it to produce new copies but still depend on the host’s ribosomes to translate their RNA into proteins to build the capsid and make new copies of their RNA polymerase. Since RNA-polymerase enzymes lack the proofreading ability of DNA-polymerase, RNA viruses mutate rapidly. A lot of human diseases are caused by RNA viruses, incluing the common cold, influenza, ebola, yellow fever, dengue fever, Zika fever, hepatitis C, rabies, polio, measles, as well as COVID-19, caused by the current apocalypse-driving coronavirus.
The Yellow fever virus is an enveloped icosahedral RNA virus.
Retroviruses, the last virus type, also have RNA as their nucleic acid. However, different from RNA viruses, retroviruses do not produce new copies directly from their RNA using a RNA-polymerase. Instead of that, they have another type of enzyme, called reverse transcriptase, that builds a DNA strain from their RNA. This viral DNA is then incorporated into the DNA of the host cell by an integrase enzyme. Retroviruses, therefore, change the host’s genome, i.e., they create a “hybrid” of themselves and the host. The infected cell then transcribes the viral DNA back into RNA, making several copies that allow to virus to reproduce. The most famous retroviruses to infect humans are Human immunodeficiency virus (HIV) and Hepatitis B virus.
Human Imunodeficiency Virus 1 is an enveloped icosahedral retrovirus that causes AIDS in humans.
But now let’s focus on our current celebrity, SARS-CoV-2, colloquially known as the coronavirus. This virus, which is causing the current apocalypse, is a new strain, discovered in late 2019, of the Severe acute respiratory syndrome-related coronavirus (SARSr-CoV). The previous SARS outbreak between 2002-2004 was caused by another strain of this same species, SARS-CoV, or now often referred to as SARS-CoV-1. This virus belongs to the genus Betacoronavirus and the family Coronaviridae. All members of the family Coronaviridae are often called “coronavirus” and the currently known species infect birds and mammals.
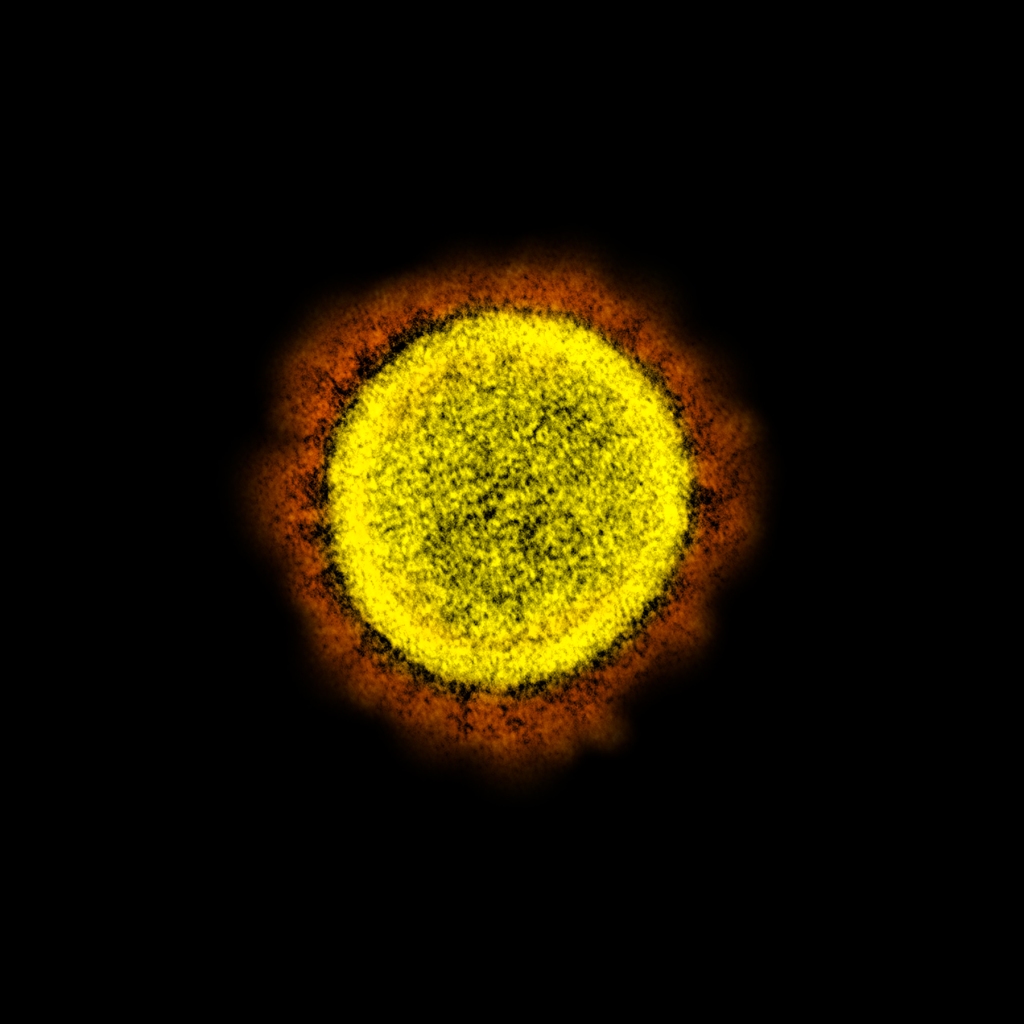
SARS-CoV-2 with artificial colors showing the “corona” (in orange) formed by the club-shaped glycoproteins of its envelope.
Coronaviruses are RNA-viruses, as mentioned above, and have a helical capsid, as well as an envelope. This envelope contains large club-shaped proteins that appear as projections on the virus surface and, in electron micrographs, resemble the solar corona, hence the name coronavirus. The envelope is built from the membrane of the host’s endoplasmic reticulum but includes glycoproteins of viral origin, including the club-shaped glycoproteins that characterize these viruses.
The presence of this envelope around the capsid has some advantages and some disadvantages to coronaviruses and any other enveloped virus. Since this envelope is basically a piece of the host’s cell, enveloped viruses can sneak into new hosts more easily because the immune system takes some time to recognize them as invaders since they are wearing a host’s “clothing”. On the other hand, this envelope is very fragile when exposed to the outer environment and degrades very quickly, so that the virus needs close contact of an infect host with a new host in order to spread. This is also why washing your hands with soap kills the virus so easily. If the virus were not enveloped, i.e., had only its capsid, it would be much more resistant.
The club-like glycoproteins of the viral envelope are also the responsible for the virus ability to infect. These glycoproteins connect to the angiotensin-converting enzyme 2 (ACE2), an enzyme that is found on the surface of many human cells. ACE2 is especially abundant in the lungs, which is the reason why this is the organ that suffers the most during SARS-CoV infections.
The main genera inside the family Coronaviridae are Alphacoronavirus, Betacoronavirus, Gammacoronavirus and Deltacoronavirus. Most known species of Alpha- and Betacoronavirus infect bats, so it is likely that the ancestor of these genera was originally a bat-specific virus that later mutated and acquired the ability to infect other species. All coronaviruses that infect humans belong to this two genera and include, besides SARS-CoV, MERS-CoV (which causes the Midle East Respiratory Syndrome) and several viruses that cause mild cold-like symptoms, such as HCoV-HKU1, HCoV-NL63 and HCoV-229E. Species of Gammacoronavirus infect mainly birds, although at least one species, Coronavirus HKU15, causes diarrhea in pigs. The genus Deltacoronavirus includes the Avian coronavirus (IBV), which causes infectious bronchitis in birds, and the Beluga whale coronavirus SW1, the only coronavirus known to infect a marine mammal.
Avian coronavirus. The club-shaped glycoproteins are clearly visible on the envelopes.
The genome of coronaviruses has about 30 thousand nucleotides, being some of the largest genomes among RNA viruses. The only known RNA virus with a larger genome, with about 41 thousand nucleotides, was discovered in 2018 and infects, guess what, planarians! Named Planarian secretory cell nidovirus (PSCNV), it belongs to the order Nidovirales, which includes coronaviruses and many other RNA-viruses, but seems to have diverged from most members of Nidovirales a long long time ago. Maybe I’ll talk more about this particular virus and the implications of its discovery in a future post.
Let’s conclude this post with a quick review of what we have learned about SARS-CoV-2, the “coronavirus”:
It is an RNA-virus, meaning that it has a great mutation potential and is able to create copies of itself in the host’s cytoplasm, being an almost self-suficient virus;
It has a helical capsid surrounding its RNA;
It has an envelope derived from the membrane of the host’s endoplasmic reticulum, which is the reason why it can be so easily killed by water and soap;
This envelope includes large clube-like glycoproteins that make it appear as a solar corona in electron micrographs, hence the name coronavirus;
It is a member of the genus Betacoronavirus, which includes a lot of species known to infect bats and that’s the reason why its origin in a Chinese bat soup is very likely.
I hope that this post helped you see more about this new virus than its ability to collapse human societies.
I’ll continue the parasite trend from last week, but this time shifting from human parasite to maize parasite, and from a prokaryotic to a eukaryotic parasite. So let’s talk about Physoderma maydis, commonly known as the brown spot of maize or brown spot of corn.
The Brown spot of maize is a fungus of the division Blastocladiomycota that infects corn (or maize) plants. Its common name comes from the fact that it causes a series of brown spots on the leaves of an infected plant.
The brown spots seen on this corn leaf are due to an infection by Physoderma maydis. Credits of the photo to Clemson University – USDA Cooperative Extension Slide Series.*
The life cycle of the brown spot of maize is as complex as that of many fungi. The infection of the plants occur through spores that remain in the soil during winter and are carried to the host by the wind, germinating in the rainy season. The germinated spores produce zoospores, flagellated spores able to swim. Swiming through the maize leaf, the zoospores infect single cells and produce zoosporangia at the surface of the leaf. The zoosporangia release new zoospores that infect new cells. In late spring and summer, the zoospores produce a thallus growing deep inside the maize leaf that infects many cells and produces thick-walled sporangia. After the plants dies and the leaves become dry and broke, the sporangia are released and reach the soil, where they wait for the next spring to restart the cycle.
The brown spot of maize is a considerable problem for maize crops in countries with abundant rainfall. Heavy infections may kill the maize plant or severely reduce its fitness before the ears are ready to be harvested. Although fungicides may help in slowing down the infectio throughout the crops, one of the most efficient ways to reduce the damage is to destroy, usually by fire, the remains of the last harvest.
– – –
References:
Olson, L. W.; Lange, L. (1978) The meiospore of Physoderma maydis. The causal agent of Physoderma disease of maize. Protoplasma 97: 275–290. https://dx.doi.org/10.1007/BF01276699
Last week I introduced a red alga, the Irish moss. Today I’m bringing another alga, this time a green one, but this is not an ordinary green alga, but a parasite of the Irish moss! So let’s talk about Acrochaete operculata, or the operculate acrochaete as I decided to call it in English, since obviously there would be no common name for an alga parasite of another alga.
Discovered and named in 1988, the operculate acrochaete is an exclusive parasite of Chondrus crispus. The infection occurs when flagellate zoospores of the parasite settle on the outer cell wall of the Irish Moss, where they start their development and digest the cell wall, penetrating the tissues of the host. In sporophytes of the Irish moss, the operculate acrochaete digests the intercellular matrix and spreads through the frond, while in gametophytes the infections remains localized, forming papules. The damages caused by the green alga lead to secondary infections by other organisms, especially bacteria, and the infected fronds end up falling apart, completely degradated.
A frond of the host (Chondrus crispus) to the left and the parasitic Acrochaete operculata that infects its tissues to the right. Photo extracted from chemgeo.uni-jena.de
As mentioned last week, the sporophytes and gametophytes of the Irish Moss have different forms of the polysaccharide carrageenan and this seems to be the reason why the parasite infects both forms differently. The sporophytes have lambda-carrageenan, which seems to increase the virulence of the parasite, while the kappa-carrageenan of the gametophyte seems to limit the green alga’s spread.
Since its discovery, the operculate acrochaete and its interaction with the Irish moss has been studied as a way to both reduce its damage on cultivated crops of the red alga and as a model to understand the relationship of plants and their pathogens.
– – –
References:
Bouarab, K.; Potin, P.; Weinberger, F.; Correa, J.; Kloareg, B. (2001) The Chondrus crispus-Acrochaete operculata host-pathogen association, a novel model in glycobiology and applied phycopathology. Journal of Applied Phycology 13(2): 185-193.
Correa, J. A.; McLachlan, J. L. (1993) Endophytic algae of Chondrus crispus (Rhodophyta). V. Fine structure of the infection by Acrochaete operculata (Chlorophyta). European Journal of Phycology 29(1): 33–47. http://dx.doi.org/10.1080/09670269400650461
Correa, J. A.; Nielsen, R.; Grund, D. W. (1988) Endophytic algae of Chondrus crispus (Rhodophyta). II. Acrochaete heteroclada sp. nov., A. operculata sp. nov., and Phaeophila dendroides (Chlorophyta). Journal of Phycology 24: 528–539. http://dx.doi.org/10.1111/j.1529-8817.1988.tb04258.x
Last week I introduced a serious plant pathogen, the gray mold, that attacks many crops and has a special role as either a bad or a good guy in wine grapes. But a plant that is never happy with an infection by the gray mold is certainly the lettuce. And in this case our juicy vegetable has an enemy that makes it susceptible to the mold, and I’m bringing it to you today.
Named Bremia lactucae, this organism is a oomycete, thus belonging to a group of organisms that was formerly classified as a fungus, but that currently is known to be more closely related to brown and golden algae. This species attacks lettuces and closely related plants, causing a disease called downy mildew.
A lettuce leaf with downy mildew. Photo by Gerald Holmes.*
The downy mildew is the most important disease affecting lettuce worldwide. The disease itself is not the main problem, although it decreases the quality of the crop. Its main problem is that it makes the vegetable more vulnerable to other infections, such as those by the gray mold, and also increases the risk of contamination by human pathogens, such as intestinal parasites.
A branch of the downy mildew under the microscope. Photo by Bruce Watt.*
The usual forms of controling the spread of the downy mildew is by using fungicides and developing mildew-resistant lettuces by hybridization with wild and naturally resistant varieties. However, as usual, the downy mildew eventually adapts to this, giving rise to fungicide-resistant strains, as well as strains able to neutralize the resistance of lettuce lineages. It’s one more evolutionary arms race.
– – –
References:
Beharav, A., Ochoa, O., & Michelmore, R. (2013). Resistance in natural populations of three wild Lactuca species from Israel to highly virulent Californian isolates of Bremia lactucae Genetic Resources and Crop Evolution, 61 (3), 603-609 DOI: 10.1007/s10722-013-0062-5
Parra, L., Maisonneuve, B., Lebeda, A., Schut, J., Christopoulou, M., Jeuken, M., McHale, L., Truco, M., Crute, I., & Michelmore, R. (2016). Rationalization of genes for resistance to Bremia lactucae in lettuce Euphytica, 210 (3), 309-326 DOI: 10.1007/s10681-016-1687-1
If I had to bet on a parasite that you who are reading this probably have in your body, I’d go for today’s fellow, the protist Toxoplasma gondii, sometimes simply called toxo.
Found worldwide, the toxo is one of the most common parasites in humans, with estimations that about half of the world’s population is infected. Fortunately, this creature usually occurs in a latent form and does not offer great risks, but eventually it may develop into a more serious condition called toxoplasmosis, especially in people with weakened immunity.
But let’s take a closer look at this tiny fellow.
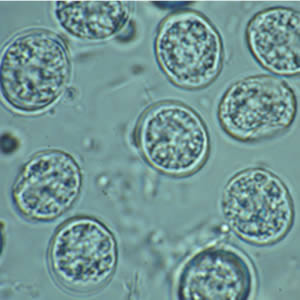
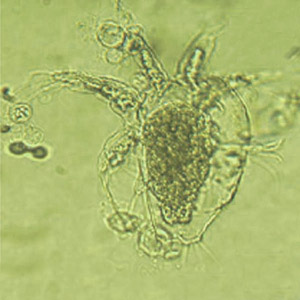
Oocysts of Toxoplasma gondii. This is the form found in the environment and that can start an infection in your body.
The toxo is a protist belonging to the phylum Apicomplexa, a group of parasitic alveolates that also includes the agent that causes malaria. Although traditionally considered a protozoan, the apicomplexans are closely related to dinoflagellates (which are generally considered as a group of algae). They have a unique organelle called apicoplast, which they use to penetrate a host cell. The apicoplast is derived from a plastid (such as the chloropast), so in a certain way we can say that the apicomplexans are algae that evolved into intracellular parasites!
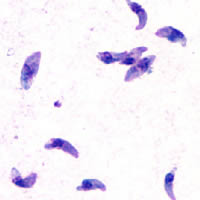
Tachyzoites of Toxoplasma gondii stained with Giesma from the peritoneal fluid of a mouse.
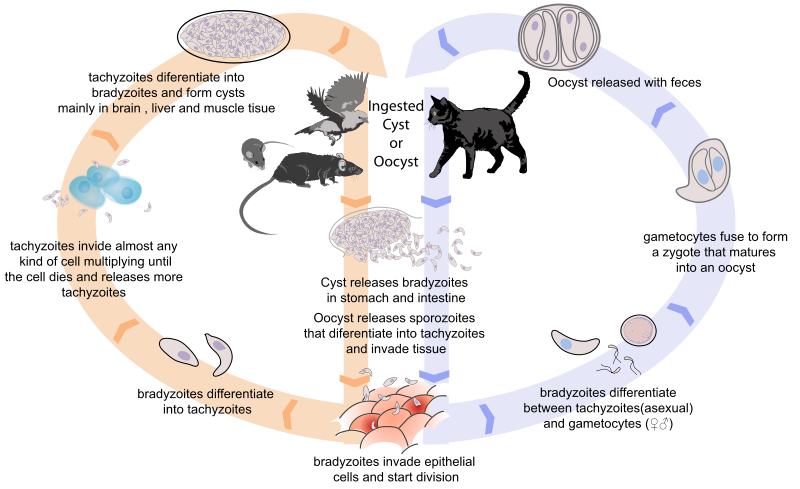
The life cycle of the toxo is kind of complex. Let’s start with the inactive form called oocyst, which may be found in the environment. If a warm-blooded animal ingests an oocyst, it will “burst” inside the gut of the animal and release several “quick-moving” forms called tachyzoites. The tachyzoites invade almost any cell of the body and multiply asexually inside it until the cell dies and release them, allowing them to infect more and more cells. When invading the brain, liver and muscles, the tachyzoites usually differentiate into cysts that become inactive. In this stage, the only thing that the toxo wants is that a cat (any species of the family Felidae) eats the host. It may even change the host’s behavior in order to make it bolder and more easily accessible to predators.
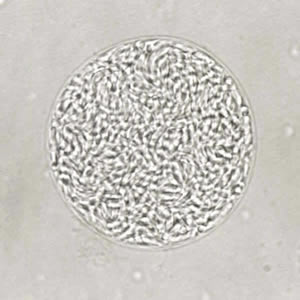
A cyst of Toxoplasma gondii that forms in the muscles, brain and liver of any warm-blooded anymal. All the cyst wants is to be eaten by a cat!
Now let’s assume that a cat ate the host (that was likely a bird or mouse). Inside the cat’s gut, the cyst burst and releases several “slow-moving” forms called bradyzoites. This form invades the epithelial cells of the cat’s intestine and multiply asexually inside them. Eventually, the bradyzoites differentiate into either tachyzoites or gametocytes (sperm- and egg-like cells). When two gametocytes fuse, they form a zygote that matures into an oocyst and is released into the environment, restarting the cycle.
The complex life cycle of Toxoplasma gondii. Credits to Mariana Ruiz Villarreal.
As always, the lifecycle of parasites is a wonderful adventure!
– – –
References:
Tenter, A., Heckeroth, A., & Weiss, L. (2000). Toxoplasma gondii: from animals to humans International Journal for Parasitology, 30 (12-13), 1217-1258 DOI: 10.1016/S0020-7519(00)00124-7
Today I’m bringing you a species that is probably one of the most terrible ones to exist today, the amphibian chytrid fungus, Batrachochytrium dendrobatidis, also known simply as Bd.
Several sporangia of Batrachochytrium dendrobatidis (spherical structures) growing on a freshwater arthropod. Photo by AJ Cann.*
The amphibian chytrid fungus, as its name says, is a chytrid, a fungus of the division Chytridiomycota, which include microscopic species that usually feed by degrading chitin, keratin in other such materials. In the case of the amphibian chytrid fungus, it infects the skin of amphibians and feeds on it. It grows through the skin forming a network of rhizoids that originate spherical sporangia that contains spores.
The infection caused by the amphibian chytrid fungus is called chytridiomycosis. It causes a series of symptoms, including reddening of the skin, lethargy, convlusions, anorexia and excessive thickening and shedding of the skin. This thickening of the skin leads to problems in taking in nutrients, releasing toxins and even breathing, eventually leading to death.
An individual of the species Atelopus limosus infected by the amphibian chytrid fungus. Photo by Brian Gratwicke.**
Since its discovery and naming in 1998, the amphibian chytrid fungus has devastated the populations of many amphibian species throughout the world. Some species, such as the golden toad and the Rabb’s fringe-limbed treefrog, were recently extinct by this terrible fungus. This whole drastic scenario is already considered one of the most severe examples of Holocene extinction. The reason for such a sudden increase in the infections is unknown, but it may be related to human impact on the environment.
We can only hope to find a way to reduce the spread of this nightmare to biodiversity.
– – –
References:
Fisher, M., Garner, T., & Walker, S. (2009). Global Emergence of Batrachochytrium dendrobatidis and Amphibian Chytridiomycosis in Space, Time, and Host Annual Review of Microbiology, 63 (1), 291-310 DOI: 10.1146/annurev.micro.091208.073435




 This work is licensed under a Creative Commons Attribution-NonCommercial 3.0 Unported License.
This work is licensed under a Creative Commons Attribution-NonCommercial 3.0 Unported License.