Polycladus gayi is an iconic land planarian found in Chile and locally known as lengua de vaca (cow tongue). Despite being the largest land planarian in Chile and one of the first land planarians to be described, back in 1845, we know almost nothing about its ecology.
However, things are changing! In the past months, two specimens have been found eating in the wild, and, in both cases, the prey was the same species, the giant black snail, Macrocyclis peruvianus, the largest snail in Chile. Both observations occurred in Chilean protected areas, Villarrica National Park and Alerce Costero National Park, and were recorded by nonspecialists visiting the areas. One more important discovery that happened thanks to citizen science!
The land planarian Polycladus gayi attached to snail Macrocyclis peruvianus and feasting on its flesh. Photo by Yerko Lloncón.*
After almost two centuries since P. gayi was discovered, we finally know something about its place in the food chain! And, of course, it also helps us see M. peruvianus from a new perspective since this also seems to be the first record of one of its predators! Even though snails are a common item in the diet of land planarians, not all species feed on them, and we cannot assume that both groups are always directly connected in the food web.
Come see how chubby the planarian got after eating the whole snail!
There is still much more to discover about these two unique Chilean creatures, and the partnership between researchers and the general public is an important way to speed up the process of gathering knowledge about the creatures around us!
Reference:
Boll PK, Lloncón Y, Almendras D (2023) Records of the land planarian Polycladus gayi (Tricladida, Geoplanidae) preying on black snails Macrocyclis peruvianus (Gastropoda, Macrocyclidae). Austral Ecology. https://doi.org/10.1111/aec.13430
We get used to thinking of the food chain, or more precisely the food web, as something considerably regular and flowing in a single direction: plants synthesize nutrients by photosynthesis, herbivores eat plants, carnivores eat herbivores and stuff. But nature does not work as regularly as we think and sometimes things can look very crazy.
Although many animals are strictly herbivores indeed, this is not exactly the case for many ungulates that we may see as vegan, such as ruminants. It is actually not that rare to find ruminants chewing on bones of dead animals, sometimes when they still have some flesh attached to them. The main reason for herbivores to eat bones is to acquire minerals, such as calcium, which are important not only for growing bones and horns or antlers but also for conducting nerve impulses and other biochemical roles.
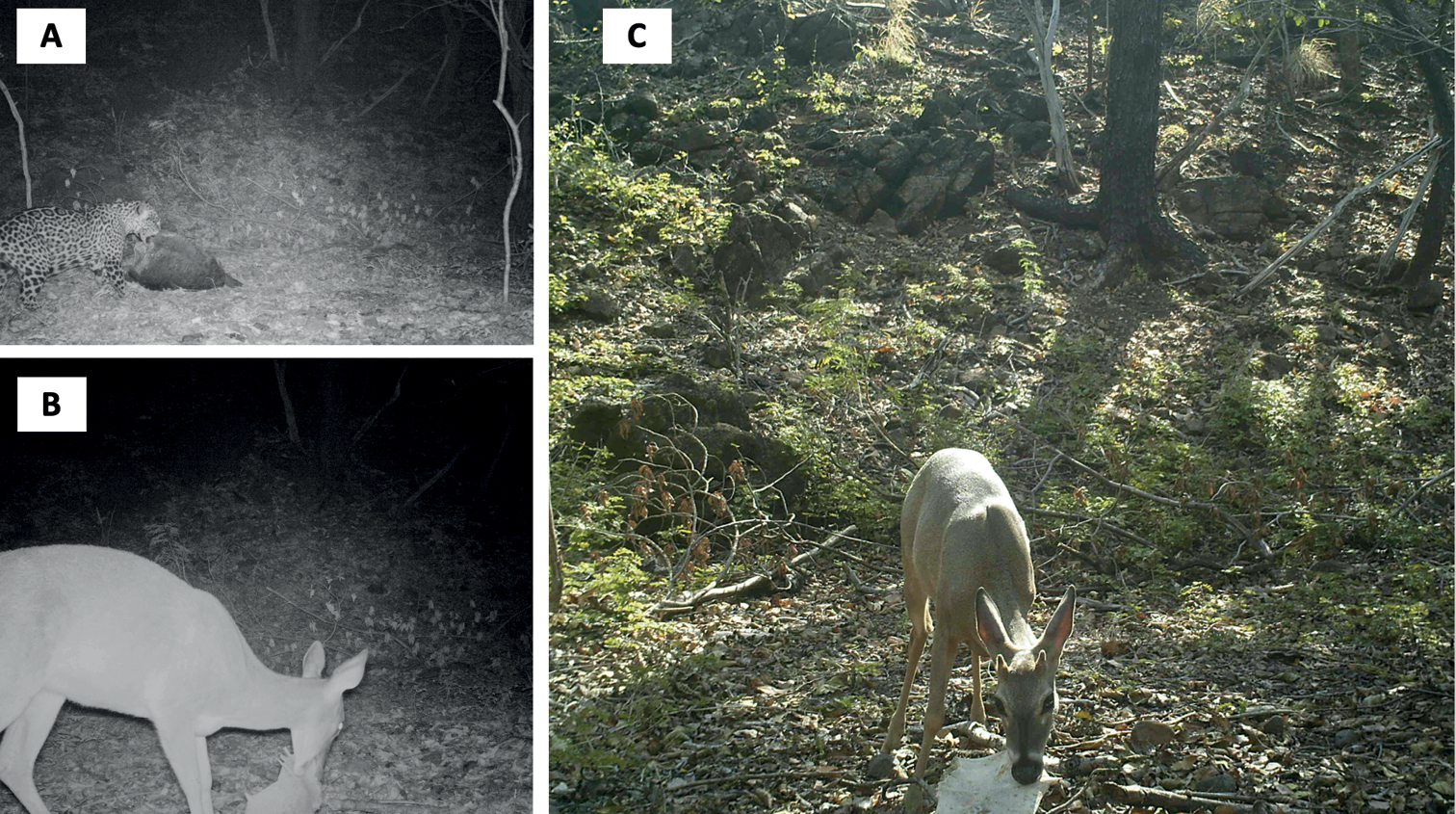
Now observations with a single camera trap in Santa Rosa National Park in Costa Rica recorded more than a hundred events of white-tailed deer (Odocoileus virginianus) chewing on bones of turtle carcasses on a beach. The deer included males with growing antlers, lactating females, and growing fawns, all of which need additional amounts of calcium to grow antlers, replace calcium lost in milk production, and grow bones, respectively.
Ok, those deer eat a lot of turtle bones, so what? Well, the reason why there are so many bones available for them to feast on is that jaguars prey on turtles that come to that beach to lay their eggs. Jaguars are among the most important predators in the Neotropics and deer are obviously one of their prey. While the presence of jaguars can be a threat to the life of the white-tailed deer, they are also essential to provide the bones on which the deer feed to get healthy.
Jaguars eat turtles on the beach and let their bones behind. Later, deer come to feed on the bones that their main predator left as a gift. “From jaguar to dear, with love, so that you can grow healthy and feed me later”. Credits to Morera et al. (2022).*
This almost turns the food web upside down, right? Or at least add some very weird loops in it.
Morera, B., Montalvo, V., Sáenz-Bolaños, C., Cruz-Díaz, J. C., Fuller, T. K., & Carrillo, E. (2022). Osteophagia of sea turtle bones by white-tailed deer (Odocoileus virginianus) in Santa Rosa National Park, northwestern Costa Rica. Neotropical Biology and Conservation, 17(2), 143-149. https://doi.org/10.3897/neotropical.17.e87274
However, in 2018, one marine polyclad seems to have decided to explore the world outside the oceans by riding a toad in Bangladesh. How did this happen? Well, no one is sure. A group of researchers working on Nijhun Dwip Island found a common Asian toad (Duttaphrynus melanosticus) walking through an agricultural field with a polyclad attached to its dorsum.
The toad and the polyclad. A new Aesop’s fable. Credits to Rabbe et al. (2020).*
A canal that passes through the field can be flooded by the sea during high tide and this is how the researchers think the flatworm ended up on land and eventually on the back of the toad. Unfortunately, because the toad was spotted by the researchers, the poor polyclad ended up collected and killed and is now preserved in a lab.
Rabbe MF, Roy DK, Mohammad N, Liza FT, Mukutmoni M, Alam MM, Begum A & Jaman MF. 2020. A Novel Natural History Phenomenon: A Free-living Marine Flatworm (Polycladida) Attached to a Common Asian Toad (Duttaphrynus melanostictus). Reptiles & Amphibians 27: 293–294. https://journals.ku.edu/reptilesandamphibians/article/view/14404
In order to try avoiding predators, many species develop powerful toxins that would harm anyone trying to eat them, sometimes even killing them. However, predators can fight back by developing a strong resistance to the prey’s defenses, sometimes to amazing levels.
Millipedes are known for as distasteful prey that evolved a variety of toxins do deter predators. Nevertheless, some species have found ways to deal with millipede’s defenses, making the poor creatures desperate for new strategies to survive.
In North America, ground beetles of the genus Promecognathus are specialist predators of millipedes. The species Xystocheir dissecta, one of their main prey, produces cyanide as a chemical defense. Cyanide is a very toxic compound for most life forms.
The cyanide-producing millipede Xystocheir dissecta. Photo by iNaturalist user mhertel.*
In a recent study, 18 different ground beetle species were exposed to sodium cyanide (NaCN) in the lab to assess their resistance. While most species succumbed in less than 10 min when exposed to 15 mg of NaCN or less, three species did not give a damn even to quantities as high as 100 mg. These three species included Promecognathus crassus, P. laevissimus and Metrius contractus. While both Promecognathus species feed on Xystocheir dissecta, Metrius contractus does not.
Promecognathus laevissimus, the “I could have cyanide for breakfast” ground beetle. Photo by Eddie Dunbar.*
In another trial, the species were exposed to 100 mg of potassium cyanide (KCN) for up to two hours. While M. contractus ramained active during the first hour, all specimens succumbed in less than two hours, but after 120 min, some specimens of Promecognathus laevissius were still moving around as if nothing was happening.
Metrius contractus, resisting cyanide just for fun. Photo by iNaturalist user tparkeressig.*
This study is the first evidence of predators having resistance to cyanide. While this superpower in P. laevissimus is easily explained by its predatory behavior, the high resistance of M. contractus is still a mystery, as this species is not specialized in millipedes, although it is possible that it may eat them as an alternative food, especially sick or injured specimens. Both species, however, are resistant to amounts of cyanide way above the ones that they would find in any millipede. It’s a real superpower.
Weary BP, Will KW (2020) The Millipede-Predation Behavior of Promecognathus and Exceptional Cyanide Tolerance in Promecognathus and Metrius (Coleoptera: Carabidae). Annals of the Entomological Society of America. https://doi.org/10.1093/aesa/saaa023
Two mosquitoes of the genus Aedes, Aedes aegypti and Aedes albopictus, are invasive species in tropical and subtropical regions worldwide. While A. aegypti is native from Africa, A. albopictus is originally from southeast Asia, but both species have been spread by humans and continue to increase their range.
Both species are known as vectors of several diseases that affect humans, especially those caused by Flavoviruses, which include the Yellow fever, Dengue fever and Zika fever. Chikungunya, caused by a species of Alphavirus is also transmitted to humans by them. Moreover, they can also transmit some nematodes, such as the heartworm that infects the heart of dogs and other carnivores.
Aedes aegypti biting a human and having a delicious bloody meal. Photo by James Gathany.
Because A. aegypti and A. albopictus pose such a huge threat to public health, getting rid of them is top priority. Here in Brazil, there is a massive national campaign to reduce the ability of Aedes to reproduce by avoiding containers with still water in the open, such as flower vases, buckets, uncovered barrels, discarded tires and virtually everything that can retain water long enough for the larvae to develop. I have to say, though, that this all seems to be useless. The mosquitoes continue to spread and the cases of dengue fever continue to grow. The fact is that the mosquitoes will find a place to lay their eggs. If they don’t find it in your backyard, they will find it in the forest or any vacant lot.
Instead of forcing them to lay their eggs where we cannot see, we should stimulate them to lay their eggs around us and then kill the larvae. Several aquatic predators have been tested as potential allies, including larvivorous fish, dragonfly nymphs, copepods, planarians and even other mosquitoes whoses larvae eat the larvae of Aedes! The use of these predators showed mixed results. Larvivorous fish are difficult to maintain in water tanks at home and dragonfly nymphs are too generalist as predators.
Now a new predator has been suggested: a plant! Yes, a carnivorous plant of the genus Utricularia, which includes species known as bladderworts. These aquatic plants have little bladder-like structures that function as traps to capture small animals. The bladder is hollow and has an internal negative pressure in relation to the environment surrounding it. This negative pressure is created by water being constanly pumped out of the bladder through its walls via active transport. The bladder’s opening is covered by a small lid that avoids water to fill it again when the trap is set. Surrounding the lid, there is a group of bristle-like protuberances. When an animal is moving through the water and moves one of those bristles, they slightly deform the lid, breaking the seal and allowing water to enter the bladder. The negative pressure then sucks water quickly into the bladder, dragging the small animal with it. Then it is only a matter of time for the poor animal to be digested.
Watch the plant in action.
A group of researchers at the University of Rhode Island, USA, tested whether Utricularia macrorhiza, the common North American bladderworth, could be an effective predator of mosquito larvae. By adding U. macrorhiza to containers with larvae of A. aegypti and A. albopictus, they were able to kill 95 to 100% of the larvae in only five days. That’s an amazing result, don’t you think?
Bladderwort with several Aedes larvae (marked with asterisks) in its traps. Credits to Couret et al. (2020).*
Since bladderworts are much easier to raise in tanks and other containers in your backyard than animal predators such as fish and dragonflies, they are a promising new alternative to control the populations of this disease-carrying insects.
So, are you eager to raise some aquatic carnivorous plants to help fight these heinous mosquitoes?
A common tropical disease in forested areas of South America and Africa is the yellow fever. Affecting most primate species, the yellow fever is usually transmitted by the famous mosquito Aedes aegypti, which also transmits the dengue and zika fevers, all caused by viruses of the genus Flavivirus.
But in forested areas of South and Central America, other mosquito species can also transmit the yellow fever to humans and monkeys. One of these species is Sabethes cyaneus, which I decided to call the blue paddled mosquito. This species occur from Mexico to Argentina and Brazil and, different from most mosquitos, is diurnal.
A female about to have a bloody lunch on a human in Mexico. Photo by Carlos Alvarez N.*
Even if you don’t find mosquitos nice creatures most of the time, you will have to admit that the blue paddled mosquito is a beautiful animal. The body of the adult is dark and has metallic blue shade on the dorsum and the legs, being slightly greener on the dorsum and slightly purpler on the legs. More than that, the second pair of legs have a large tuft of hair that makes it look like a pair of paddles.
But what is the function of those paddles? The first guess would be that they are sexually selected and are likely important during courtship behavior. But females also have paddles and, if they were the result of sexual selection caused by females on males, they would likely be much larger on males, which is not the case.
Another female feeding on a human, this time in Paraguay. Photo by Joaquin Movia.*
Males perform, indeed, a complex courtship ritual in front of the females using their paddled legs. When females are prepared to mate, they perch vertically on a branch and wait for males to come and dance before them. Most of the males are rejected by a female and, when she finally chooses a male, she will compulate only with him. Males, on the other hand, copulate with many females. This increases even more the idea that the paddles must have some importance on female choice.
This is not what was found, though. When the paddles of a male are reduced in size or removed completely, he still has the same chances of getting a female than intact males. On the other hand, a female whose paddles were removed rarely attracts any male. She remains perched on her branch waiting and waiting and no male will come to dance for her. The interest that male blue paddled mosquitos have for paddles is so strong that they even approach other perched males with large paddles.
The reason why this species exhibits strong male preference and weak female preference is still a mystery but is a nice reminder that our ideas on sexual selection are not as well-established as we might think.
Hancock RG, Foster WA, Lee WL (1990) Courtship behavior of the mosquito Sabethes cyaneus (Diptera: Culicidae). Journal of Insect Behavior 3(3): 401–416. doi: 10.1007/BF01052117
South SH, Arnqvist G (2008) Evidence of monandry in a mosquito (Sabethes cyaneus) with elaborate ornaments in both sexes. Journal of Insect Behavior 21: 451. doi: 10.1007/s10905-008-9137-0
South SH, Arnqvist G (2011) Male, but not female, preference for an ornament expressed in both sexes of the polygynous mosquito Sabethes cyaneus. Animal Behaviour 81(3): 645–651. doi: 10.1016/j.anbehav.2010.12.014
During the past three weeks, I presented a fig tree, the Chinese Banyan, a thrips that parasitizes it, the Cuban Laurel Thrips, and a mite that parasitizes the thrips, the Cuban-Laurel-Thrips Mite. However, I haven’t wrote yet about one of the most interesting creatures that interacts with a fig tree: its pollinator.
In the case of the Chinese Banyan, its pollinator is the fig wasp Eupristina verticillata, which I named the Chinese Banyan Wasp. As all fig wasps, this species is very small and completely adapted to live with figs. They cannot survive without the exact fig species with which they interact and the fig species cannot reproduce without that exact wasp. How does this works?
Let’s start our story with an adult female Chinese banyan wasp. The females are black and very small, measuring around 1 to 1.2 mm in length only. This female is flying around looking for a young fig which will serve as her nest and her grave.
This is what a female Chinese Banyan wasp loooks like. Photo by Forest & Kim Starr.*
A fig, in case you don’t know, is not a real fruit in the botanical sense. It is actually a special kind of inflorescence called a syconium that is basically a flower-filled sack. The inner walls of a fig have many tiny male and female flowers and the only way to get to them is through a tiny hole at the fig’s appex. And this hole is only open during the initial stages of the fig’s development.
Chinese Banyan figs in their early stage. You can see the hole marked by a darker “areola” around them. That is the place through which a female fig wasp enters the fig. Credits to Wikimedia user Vinayaraj.**
When the female Chinese Banyan fig wasps is flying around, she is looking for a fig that is at this exactly stage of development. Once she finds one, she crawls inside the fig through that tiny hole. She usually loses her wings while doing that because the passage is too narrow. She evens needs to use her especially adapted mandible to help her go through. Once inside the fig, she looks for the female flowers, which are located at the base of the fig, away from the entrance. The male flowers, located right at the entrance, are not mature yet. However, the female wasps arrived with pollen that she gathered elsewhere (you will learn about that soon). When she reaches the female flowers, she introduces her ovopositor (the long structure at the end of her abdomen that is used to lay eggs) inside the female flower and lays one egg inside the flower’s ovary. Her ovopositor needs to have the exact size to reach the ovary to lay the egg. If it is too short, she is unable to complete her task. And while she is moving from flower to flower to lay eggs, she ends up pollinating them. After she has finished, she dies still inside the fig.
The ovaries that received an egg start to grow into a gall (a “plant tumor”) by influence of the insect and serve as food and shelter for the larvae that hatch from the eggs. A larva grows, pupates and turns into an adult inside a single gall. When the wasps have finally reached their adult stage, they leave the gall in which they were born. This happens when the fig reached its mature stage.
Males are the first ones to emerge. They are even smaller than the females and have a yellow to light-brown color. They gnaw their way through the gall and, once outside it (but still inside the fig) they start to look desperately for female wasps to inseminate. They do that by tearing other galls apart and, when a female is found trapped inside, they inseminate her. After that, the males dig a hole through the fig to the outside and die soon after, never experienced the external world.
A male Chinese Banyan wasp (right) compared to a female. Photo by Forest & Kim Starr.*
Female wasps then leave their galls and move towards the hole opened by the male. While doing that, they move over the now mature male flowers and become covered in polen. After leaving the fig, they search for another fig that is in its early stage of development, restarting the cycle.
When a female leaves a mature fruit, she needs to find an immature one soon after that because she will die in a couple of days. In other words, the only way for this to work is if there are figs in the right stage all year around, and that is what happens. Differently from most plant species, which produce flowers in a specific time of the year, fig trees are always flowering. Well, not exactly. One individual fig tree produces figs only in a specific period of the year. All the figs of that tree ripen at the same time, i.e., a fig tree has an intra-individual synchrony of flower maturation. However, other trees of the same species have different moments to produce flowers, i.e., there is an inter-individual asynchrony of flower maturation. This assures that a wasp will always find a fig at the suitable maturation stage when there are enough fig trees around and also assures that a fig tree will not be fertilized by its own pollen.
As I mentioned when I presented the Chinese Banyan, this tree can only produce viable figs when the wasp is present, so that populations introduced outside of their native range will only reproduce if the waps is introduced as well. However, the wasp will be unable to survive if there are not enough fig trees to provide it with figs all year round. It is a delicate relationship between a tiny, fragile and short-lived insect and a huge, resistant and long-lived tree. And they need each other to survive.
McPherson JR (2005) A Recent Expansion of its Queensland Range by Eupristina verticillata, Waterston (Hymenoptera, Agaonidae, Agaoninae), the Pollinator of Ficus microcarpa l.f. (Moraceae). Proceedings of the Linnean Society of New South Wales: 126: 197–201.
Weiblen DG (2002) How to be a fig wasp. Annual Review of Entomology 47: 299–330.
I recently presented a thrips in the Friday Fellow section, in that case a thrips that infects mostly fig trees. This group of insects, which make up the insect order Thysanoptera, is poorly known by the general public, but is certainly known by gardeners and farmers, as they can be a serious nuisance for many plant types.
We could imagine thrips as being kind of the mosquitoes of plants. They pierce the surface of plants and suck their juices just like mosquitoes do with vertebrates. And we all know that a mosquito bite may lead to much more than a small blood loss and local irritation of the skin. Many parasites use mosquitoes as vectors to travel from host to host, including protists such as Plasmodium falciparum, which causes malaria, and many types of virus, such as those of the genus Flavivirus, which cause the yellow, dengue and zika fevers.
A similar thing happens in the association of thrips with plants. A special genus of virus, called Tospovirus, infects many plant species and uses thrips as a vector. Inside the thrips bodies, the viruses reproduce after infecting the epithelial cells of the gut and, from there, travel via blood to the salivary glands and, when a thrips perforates a plant, the virus is injected in it. The cycle is basically the same used by Flavivirus in mosquitoes and ticks to infect vertebrates. Isn’t it amazing how a virus such as Tospovirus can infect both an animal and a plant? But what exactly is the disease caused by these viruses?
Basil leaf infected with the tomato spotted wilt virus. Photo by Scot Nelson.**
One of the most common Tospovirus is the so-called Tomato spotted wilt virus (TSWV), which is considered one of the most economically devastating plant viruses in the world. It can infect many crops, such as tomato, tobacco, bellpepper, peanut and basil. The symptoms vary from plant to plant, but usually include stunting, poorly developed fruits, commonly with ring spots on the surface, and necrosis of the leaves. It is transmitted to plants by thrips of the genus Frankliniella, mainly the western flower thrips Frankliniella occidentalis. Although the virus usually needs several hours to be able to reinfect a plant after infecting a thrips, in ideal conditions the time can e as short as five minutes.
The western flower thrips Frankliniella occidentalis. Photo by Dave Kirkeby.*
But why would a thrips feed on an obviously sick plant, all ugly and full of spots? They would certainly prefer a healthy plant, but that would prevent the virus to spread. As a result, the virus developed several strategies to attract the thrips. The TSWV is able to increase the amount of free aminoacids in infected plants, and these are essential nutrients for egg production in thrips. As a consequence, infected plants become more nutritious and attract more thrips. Feeding on infected plants, the thrips will certainly get infected and at the same time ingest more nutrients than non-infected thrips. Thus, a sick thrips actually has an increased fitness and usually lays more eggs. The plants would certainly get effing scared if they were able to have emotions.
The Soybean vein necrosis virus (SVNV) is another Tospovirus of economic concern. As it names suggests, it attacks mainly soy plants, and its main vector is the soybean thrips Neohydatothrips variabilis. Infected soybean thrips produce significantly more offspring than non-infected ones, although heavily infected individuals lay few viable eggs. How do thrips bypass this problem? It’s simple! Once they are infected, they stop feeding on infected plants and prefer non-infected ones, which increases their reproductive success by avoiding becoming heavily infected and at the same time they spread the virus further to non-infected plants. A nightmare for the plants once more.
Soybean thrips Hydatothrips variabilis. Photo by Even Dankowicz.***
A recent study investigated the relationship of another Tospovirus-thrips pair, this time of the iris yellow spot virus (IYSV), which commonly attacks garlic and onion plants, and its main vector, the onion thrips, Thrips tabaci. Infected thrips did not show an increased daily fecundity but had an increased lifespan, allowing them to lay more eggs simply because they lived longer.
But the effect of Tospovirus on thrips can go further. For example, although plants infected by the TSWV release more aminoacids that attract and increase the fecundity of thrips, the infections still seems to have some deleterious effects on the insect. Infected males of Frankliniella occidentalis increase their consumption of food juices and increase the transmission of the virus. Females, on the other hand, seem to need nutrients that cannot be found in plants. As a result, they increase the consumption of eggs of the two-spotted spider mite Tetranychus urticae, with which they often coexist. Although primarily herbivorous as most thrips, the western flower thrips eventually feeds on mite eggs, and being infected by TSWV makes females become more eager to eat eggs. This is certainly not a strategy of the virus itself as the other ones, since a female that is feeding on mite eggs does not contribute for the virus’ reproductive success. Nevertheless, this is an interesting phenomenon that show us how the interactions in a trophic web can be dynamic, changing, for example, due to an uninentional side effect of a virus trying to survive.
Keough S, Han J, Shuman T, Wise K, Nachappa P (2016) Effects of Soybean Vein Necrosis Virus on Life History and Host Preference of Its Vector, Neohydatothrips variabilis , and Evaluation of Vector Status of Frankliniella tritici and Frankliniella fusca. Journal of Economic Entomology 109(5): 1979–1987. doi: 10.1093/jee/tow145
Leach A, Fuchs M, Harding R, Nault BA (2019) Iris Yellow Spot Virus Prolongs the Adult Lifespan of Its Primary Vector, Onion Thrips (Thrips tabaci) (Thysanoptera: Thripidae). Journal of Insect Science 19(3): 8. doi: 10.1093/jisesa/iez041
Shrestha A, Srinivasan R, Riley DG, Culbreath AK (2012) Direct and indirect effects of a thrips‐transmitted Tospovirus on the preference and fitness of its vector, Frankliniella fusca. Entomologia Experimentalis et Applicata 145(3): 260–271. doi: 10.1111/eea.12011
Stafford-Banks CA, Yang LH, McMunn MS, Ullman DE (2014) Virus infection alters the predatory behavior of an omnivorous vector. Oikos 123(11): 1384–1390. doi: 10.1111/oik.01148
When we think of animals changing colors to adapt to the background, we readily think of chameleons, or maybe of some extremely rapid color switchers such as cephalopods like octopuses and cuttlefish. However, many other animals have this ability too.
One example are tree frogs of the family Rhacophoridae, especially of the genus Rhacophorus.
Recently, the phenomenon was recorded for the first time for the species Rhacophorus smaragdinus in northeastern India. The animal was of a vivid green color when found but, as soon as the researchers handled it, it turned into a dull brown color in a matter of seconds, only to slowly go back to green after left alone.
A disturbed female of Rhacophorus smaragdinus made itself brown and started to get green again after left undisturbed for some minutes. Credits to CK et al. (2019).*
CK D, Payra A, Tripathy B, Chandra K (2019) Observation on rapid physiological color change in Giant tree frog Rhacophorus smaragdinus (Blyth, 1852) from Namdapha Tiger Reserve, Arunachal Pradesh, India. Herpetozoa 32: 95–99. doi: 10.3897/herpetozoa.32.e36023
Sexual cannibalism is the act of eating a sexual partner right before, during or right after copulation. Despite being a considerably rare behavior, its occurrence is very popular among the general public.
When sexual cannibalism occurs, it usually consists of the female eating the male. Two popular cases are those of mantises and of spiders, especially the black widow. This phenomenon, at least among black widows, is much rarer than most people think.
Female mantis eating a yummy male. Photo by Oliver Koemmerling.*
Although sometimes sexual cannibalism occurs because one of the partners mistakes the other for food, in many species it is a evolutionary selected strategy that assures that the female will eat enough for the offspring to develop properly. It may look horrible from our human point of view, especially if we think from the perspective of the male, but we must remember that passing your genes to the next generation is the main purpose of most organisms and, if the male succeeded in fertilizing the female’s eggs, his life has served his purpose and he can die happily.
Sexual cannibalism is, of course, almost exclusively observed among predators, which is kind of obvious. And, as I said above, is commonly performed by the female. One group that is famous for its female-empowered species is the insect order Hymenoptera, which includes bees, ants, wasps, sawflies, among others. Since many hymenopterans have some degree of sociality, in which societies are composed almost exclusively of females, and males are generated only for the purpose of reproduction, it is curious that sexual cannibalism has never been recorded in this group… until now.
A recently published study examined the mating behavior of a small parasitoid wasp, Gonatopus chilensis. This species belongs to the family Dryinidae, of which all species lay their eggs on insects of the suborder Auchenorrhyncha, which includes cicadas, leaf hoppers, plant hoppers, among others. The larvae, after hatching from the egg, feed on the hosts. Adult females of dryinid wasps are also voracious predators and feed on the same species on which they fed as larvae.
Male of Gonatopus chilensis (left) inseminating a female (a), and female eating a male (b and c). Extracted from Virla & Espinosa (2019).*
After copulation, females of G. chilensis were often observed trying to capture the males in the same way they capture their prey. However, in only one occasion the female was successful in capturing the male and ate its gaster (the large round portion that forms most of the abdomen in wasps). Since only one instance of cannibalism was observed, it may be a rare phenomenon in this species, but since several attempts to capture the male were seen, it seems that eating the male is an interesting idea for the females.
This is the first known case of sexual cannibalism in hymenopterans and, therefore, an important record that increased the number of groups in which this behavior is known to occur.
Virla EG, Espinosa MS (2019) Observations on the mating behavior of a dryinid and first record of sexual cannibalism in the hymenoptera. Acta Ethologica. doi: 10.1007/s10211-019-00315-9


 This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.