The world is filled with bacteria. Trillions, quadrillions, quintillions of them all around us. But while most of them are harmless or even beneficial to us, others are really evil.
Currently, one of the most dangerous bacteria for humans is called Pseudomonas aeruginosa, which I decided to nickname the green-pus bacterium and you will soon find out why. With a typical rod shape as many bacteria, its cells measure about 1.5 to 3 µm in length and 0.5 to 0.8 µm in width.
Gram-sainted cells of Pseudomonas aeruginosa under the microscope. Photo by Wikimedia user Y_tambe.**
Find all around the world, the green-pus bacterium is one of the most generalist species that we have ever discovered. It often lives in oxygen-rich environments, but can also survive in anaerobic places. Provided that the environment has some moisture, it can thrive anywhere, including soil, water, on the surface of animals, plants, and fungi, and on the surface of human-made objects.
The green-pus bacterium feeds on almost anything organic, including living tissue and even hydrocarbons. It has been used, for example, to clean soil and water after oil spills by eating away the oil.
Although not exactly a parasitic species, the opportunistic and generalist habits of the green-pus bacterium make it a potential pathogenic species for humans and many other organisms. If it has the chance to feed on live tissue, it will. And this is not as difficult to occur, as this species can be found living on our skin as part of our microbiota. And how wouldn’t it be there, right? As I said, it thrives everywhere.
Nevertheless, this species is mostly harmless in normal conditions for healthy individuals. It is especially dangerous to immunocompromised individuals or other seriously injured ones. It is one of the most common species to spread and cause hospital-acquired infections. Carried through the air, it can infect the respiratory tract of immunocompromised people and cause pneumonia. Similarly, it can penetrate the urinary tract through infected catheters and cause urinary infections or, also through catheters, it can end up in the bloodstream. Another important route to infect humans is through severe skin burns, through which it can reach the inner tissues and spread, eating the infected individual alive.
Infections by the green-pus bacterium produce, as you may have guessed, a characteristic green pus. When cultivated in the laboratory, the cultures also show a blue-green color, hence the epithet aeruginosa, which in Latin means verdigris-colored. This color is caused by two metabolites produced by the green-pus bacterium named pyocyanin (with a blue color) and pyoverdine (with a green color).
Cultures of P. aeruginosa often show a greenish color. Photo by Wikimedia user Sun14916.**
One of the most frightening facts about the green-pus bacterium is that it is resistant to most antibiotics, most of which is due to natural resistance, although it also easily acquires new resistances by natural selection after being exposed to them during antibiotic treatments. This makes it very difficult to fight an infection caused by this species, and an infected person can easily die. As a result, the green-pus bacterium is also one of the most studied bacteria in the world and it stimulates research for the development of new methods to fight against bacterial infections that go beyond the use of antibiotics.
So take care! This little fellow is all around us and, although harmless most of the time, it will not hesitate to infect you if a good opportunity arises.
Diggle, S. P., & Whiteley, M. (2020). Microbe Profile: Pseudomonas aeruginosa: opportunistic pathogen and lab rat. Microbiology, 166(1), 30. https://doi.org/10.1099/mic.0.000860
It’s time for another bacterium and why not one that loves us even though we hate it? Chlamydia trichomatis is today’s fellow, that little annoying bacterium that infects us humans and sometimes can cause us some serious problems.
The human chlamydia belongs to a phylum of bacteria known as Chlamydiae, which is simply the plural of the bacterium’s name. All Chlamydiae seem to be obligate endosymbionts of eukaryotic cells, either as parasites or in a mutualistic relationship. The human chlamydia, of course, is of the first type. This species is an exclusive parasite of humans and apparently cannot infect the cells of any other species.
The life cycle of the human chlamydia is similar to that of other chlamydia species. It has two distinct forms known as elementary bodies and reticulate bodies. The elementary bodies are a spore-like form measuring from 200 to 400 nanometers in diameter. They have a very rigid cell wall and are able to survive outside of a host cell. When an elementary body contacts a human host cell, mostly cells from the mucous membranes, it causes the host cell to make a vacuole in which it remains. This vacuole is known as an inclusion.
Chlamydia inclusions (the large bubbles in the central cell) as seen under the microscope.
Within the inclusion, the elementary body changes into the metabolically active reticulate body, which measures between 600 and 1500 nanometers. The reticulate body is capable to change the inclusion into a more suitable environment and starts to replicate very rapidly until filling the host cell with bacteria in up to 72 hours. At this point, the reticulate bodies change back to elementary bodies and make the host cell burst and release them, where they can spread to other cells and infect them as well.
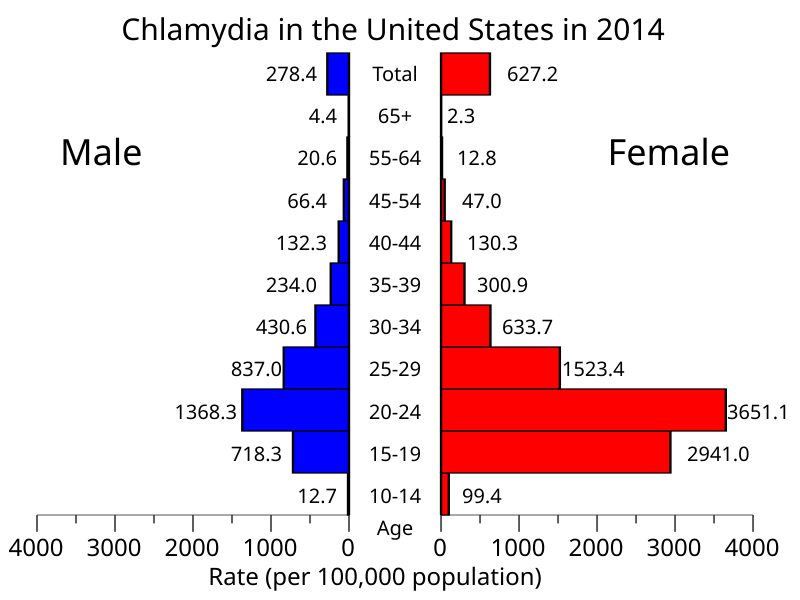
The human chlamydia can infect many parts of the human body, but the most commonly affected areas are the urethra and the vagina and its transmission between humans occurs mainly through sexual intercourse as the infected person can have elementary bodies in its fluids, such as sperm and vaginal fluid. In fact, chlamydia is the most common sexually transmitted infection worldwide, with about 4.2% of all women and about 2.7% of all men having it.
Many cases of chlamydia infection can go undiagnosed because sometimes the infection does not cause any symptoms or they take a long time to appear. When it infects the vagina and cervix, symptoms are rare at first but as the infection spreads it can infect the rest of the reproductive system and cause pelvic inflammatory disease, which may lead to sterility. Some of the rare symptoms in a vaginal infection are pain during intercourse and vaginal bleeding. In the urethra, symptoms are more common and include pain or a burning sensation during urination and eventually an unusual discharge from the penis. The symptoms are very similar to those of gonorrhea.
Besides the urogenital tract, the human chlamydia can infect many other sites, such as the rectum and the oral cavity through anal and oral sex, respectively. Another commonly infected area are the eyes, with 19% of all cases of conjunctivitis being caused by the human chlamydia. If not adequately treated, this conjunctivitis evolves into a chronic condition known as trachoma that often leads to blindness. The eyes become infected by direct contact of infected hands or objects (such as towels) with the eyes. The bacterium can also be transmitted by flies as they move around human bodies licking their fluids.
Chlamydia infection by age and sex in the United States.
Chlamydia is often treated by antibiotics such as azithromycin. Prevention includes adequate hygiene, safe sexual practices and regular testing in sexually active humans since identifying the infection earlier can reduce its damage and prevent its spread to others. In the past decade, there has been an increased interest in developing a vaccine against chlamydia. One problem is the fact that the immune response against this bacterium seems to be very complex. However, preliminary tests with a candidate vaccine has led to promising results, so there is hope!
Despite being a pain in the ass (or most commonly in the crotch) for humans, the human chlamydia is at the same time a fascinating organism just like every other lifeform on Earth. It has a considerably small genome, with only bout 900 genes. Many essential metabolic genes are lacking and it is believed that they are scavenged from the host.
A human-specific parasite, the human chlamydia is believed to have become a separate lineage from other chlamydia species about 9 million years ago. This means it has been with us since before we were even humans.
Brunham, R. C., & Rey-Ladino, J. (2005). Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nature reviews immunology, 5(2), 149-161. https://doi.org/10.1038/nri1551
Manavi, K. (2006). A review on infection with Chlamydia trachomatis. Best Practice & Research Clinical Obstetrics & Gynaecology, 20(6), 941-951. https://doi.org/10.1016/j.bpobgyn.2006.06.003
Bacteria are some of the most diverse organisms on Earth and they perform all sort of ecological roles, although they are more often associated with diseases by the average human being. This is, of course, due to the fact that most bacteria that have a direct and perceptible influence on human life are, in fact, pathogenic, often parasitic, bacteria. Today I am introducing one of those pathogenic bacteria, but not one that infects humans.
Currently known as Flavobacterium psychrophilum, this species is known to infect freshwater fish, causing a disease known as bacterial coldwater disease (CWD). As a result, I will call this species the coldwater disease bacterium, or CWD bacterium for short.
The typical aspect of CWD bacteria seen under the microscope. Extracted from Cipriano & Holt (2005).
The CWD bacterium is a typical rod-shaped bacterium measuring up to 1 µm in width and 5 µm in length. It lacks any type of flagellum or other structure that helps it move, but it can move by gliding, although this movement is incredibly slow and very difficult to observe. When cultured in a growth medium, they produce small 3-mm-diameter yellow colonies with thin margins.
Several colonies of CDW bacteria on a culture medium. Credits to Eva Säker (SVA) & Karl-Erik Johansson (BVF, SLU & SVA).*
Living in freshwater, the CWD bacterium prefers cold waters, with temperatures of 16 °C or lower, with the optimal temperature being 13°C. They grow on all sort of tissues on the body of fish, such as the skin, gills, mucous and internal organs such as the brain, kidney, spleen and the sex organs. Its preferred hosts are salmonoid fishes, such as salmons and trouts, but it can be found in other species eventually.
The CWD bacterium is an aerobic bacterium but is unable to use carbohydrates as a source of carbon, feeding on peptides instead. Thus, it secretes enzymes on the host’s tissues to degrade its proteins into peptides, causing structural damage.
Infected fish show tissue erosion, which often begins on the caudal fin and eventually spread. Fins become dark, ragged, split or torn and may be completely lost. Ulcerations appear on the skin, especially around the jaw, and the fish present behavioral issues such as spiral swimming and lethargy. The infection often kills the fish but sometimes a milder chronic infection can occur, which, however, still causes considerable behavioral changes in the host.
Lesions caused by Flavobacterium psychrophilum in the rainbow trout Oncorhynchus mykiss (A) and the coho salmon Oncorhynchus kisutsch. Extracted from Starliper et al. (2011).
The bacteria are often transmitted from fish to fish via direct fish contact, but infected adult fish can end up passing the infection directly to their offspring through infected eggs. The infection can be treated in early stages using the antibiotic oxytetracline or by adding quaternary ammonium cations to the water.
In natural environments the problems caused by this infection are rarely problematic and its damage is more often seen in fish farms, where the poor creatures are forced to live in higher densities, which increases the bacterium’s success. Apparently native to North America, where it was discovered in the 1940s, it was spread via fish farming across the whole world in the following decades. We humans, therefore, are once more the main reason why this species has become a worldwide problem.
– – –
References:
Cipriano, R. C., & Holt, R. A. (2005). Flavobacterium psychrophilum, cause of bacterial cold-water disease and rainbow trout fry syndrome. Kearneysville, WV: US Department of the Interior, US Geological Survey, National Fish Health Research Laboratory.
Langevin, C., Blanco, M., Martin, S. A., Jouneau, L., Bernardet, J. F., Houel, A., … & Boudinot, P. (2012). Transcriptional responses of resistant and susceptible fish clones to the bacterial pathogen Flavobacterium psychrophilum. PLoS One, 7(6), e39126. https://doi.org/10.1371/journal.pone.0039126
Starliper, C. E. (2011). Bacterial coldwater disease of fishes caused by Flavobacterium psychrophilum. Journal of Advanced Research, 2(2), 97-108. https://doi.org/10.1016/j.jare.2010.04.001
Bacteria are found almost everywhere across our planet and they are essential for the survival of every other lifeform, including the fascinating, and for some disgusting, cockroaches. One special cockroach-friendly genus of bacteria has the adequate name Blattabacterium, whose best-known species is Blattabacterium cuenoti, which I decided to call the “common cockroach bacterium”.
This interesting species, like all species of Blattabacterium, is an obligate endosymbiont of cockroaches, meaning that it can only exists inside cockroach cells. More specifically, the common cockroach bacterium lives inside the cells of the fat bodies of cockroaches, i.e., their adipose tissue. It was found living inside all cockroach species examined to date with the exception of the genus Nocticola. It is also found inside the termite Mastotermes darwinensis because, if you did not know yet, termites are nothing more than highly specialized cockroaches. Thus, it is thought that this bacterium first “infected” the ancestor of all modern cockroaches about 140 million years ago and has only been lost in two lineages, the one from Nocticola and the one from termites.
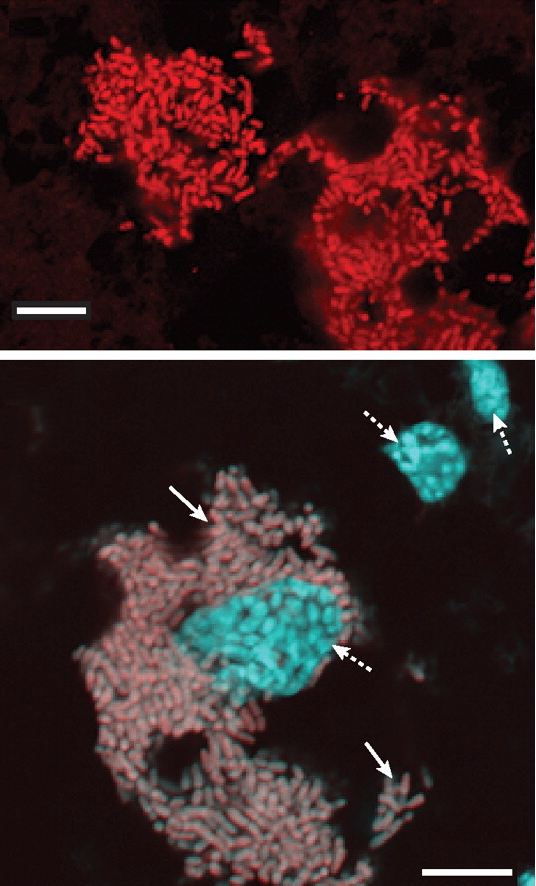
Blattabacterium cuenoti cells shown in red (above) and gray (below). The cyan areas in the bottom image represent the nucleus of the cockroach cells. Extracted from Sabree et al. (2009).
Although many cockroaches are generalist feeders, being able to feed on almost everything, the main diet of all species is decaying plant material, and this is a relatively nitrogen-poor food. In order to increase their nitrogen intake, cockroaches store uric acid, a common product of protein metabolism. Most animals, including humans, excrete uric acid in their urine, but cockroaches store it in their adipose tissue. Thus, it was originally thought that the cockroach bacteria, by living close to uric acid reserves in the adipose tissue, could use uric acid directly as a food source, but studies have found this is not the case.
When necessary, cockroaches release this uric acid and it is broken down into urea or ammonia by bacteria living in their guts. After that, the common cockroach bacteria can use those compounds to synthesize glutamate, essential amino acids and vitamins for the cockroach.
Since they cannot use uric acid directly, it is a mystery why the common cockroach bacteria lives so close to the place where this substance is stored. One suggestion is that it was originally able to use uric acid but lost this ability by genome reduction.
The functional gene categories of Blattobacterium are very similar to those of Blochmannia, an endosymbiotic bacterium from carpenter ants, which also feed on plant material. However, Blochmannia is very distantly related to Blattobacterium, suggesting that their similar genomes are the result of convergent evolution caused by similar lifestyles.
When something works, nature invents it more than once.
López-Sanchez MJ, Neef A, Peretó J, Patiño-Navarrete R, Pignatelli M, Latorre A, Moya A (2009) Evolutionary Convergence and Nitrogen Metabolism in Blattabacterium strain Bge, Primary Endosymbiont of the Cockroach Blattella germanica. PLoS Genetics 5(11): e1000721. 10.1371/journal.pgen.1000721
Patiño-Navarrete R, Moya A, Latorre A, Peretó J (2013) Comparative Genomics of Blattabacterium cuenoti: The Frozen Legacy of an Ancient Endosymbiont Genome. Genome Biology and Evolution 5(2): 351–361. https://doi.org/10.1093/gbe/evt011
Sabree ZL, Kambhapati S, Moran NA (2009) Nitrogen recycling and nutritional provisioning by Blattabacterium, the cockroach endosymbiont. PNAS 106(46): 19521–19256. https://doi.org/10.1073/pnas.0907504106
There are more than 3000 species of cicada worldwide and they are often associated with the summer when they become adults and their songs can be heard coming from the trees. Today we will focus on an Australian species, Psaltoda claripennis, known as the clanger cicada.
The clanger cicada is found in eastern Australia and is common around in Brisbane and nearby areas, where it can be easily seen on tree branches, sometimes in groups. They have a brownish dorsum with some dark, sometimes black, segments in the abdomen. The ventral side is pale, except for the abdomen, which is brown, and the legs are yellow. The eyes are light red to brownish red and the wings are transparent with green veins. Males measure about 30 mm in length and females are slightly smaller, about 25 mm long.
Male (left) and female (right) clanger cicada in Brisbane, Australia. Extracted from brisbaneinsects.com
I wasn’t able to find much information about its natural history. This species was actually just one more among many cicada species until some years ago when an interesting discovery was made.
Cicada wings are beautiful structures and are usually very clean. In fact, many insect species find ways to maintain their wings clean even in very contaminated environments and one of the reasons for it is that insects wings are extremely hydrophobic, i.e, they repel water just like many plant leaves. Since water has a hard time trying to attach to their wings, microorganisms associated with water cannot get to the wings either.
But the wings of the clanger cicada are more than only hydrophobic. Studies have shown that every cell of gram-negative bacteria that happens to touch the wing surface is deformed and dies. The same did not happen with gram-positive bacteria, though. As the studies progressed, researches started to understand the structural arrangement of the wings. Their surface is formed by very small pillars, only about 30 nm high and distant 170 nm from each other. When a gram-negative bacterium falls on those pillars, its soft membranes start to slide to the space between them and stretch enough to break. The poor cell ends up as a dead disformed mass. Gram-positive bacteria have more rigid cell walls and are resistant to stretch, but treating them with microwave decreased their rigidity and allowed them to be killed as well.
Nanostructure of the clanger cicada’s wing and the representation of how a bacterium dies by touching it. Credits to Pogodin et al. (2013).
Further research on this structure can lead to the development of new materials that remain sterile even after contacting a pathogen.
Once more the diversity of lifeforms brought us ways to improve our society. How many more useful stuff are hidden in the forests and fields? Preserving the ecosystems is the best for every inhabitant of this planet.
Xue F, Liu J, Guo L, Zhang L, Li Q (2015) Theoretical study on the bactericidal nature of nanopatterned surfaces. Journal of Theoretical Biology 385:1–7. https://doi.org/10.1016/j.jtbi.2015.08.011
Hasan J, Webb HK, Truong VK et al. (2013) Selective bactericidal activity of nanopatterned superhydrophobic cicada Psaltoda claripennis wing surfaces. Applied Microbiology and Biotechnologt 97:9257–9262. https://doi.org/10.1007/s00253-012-4628-5
Pogodin S, Hasan J, Baulin VA et.al. (2013) Biophysical Model of Bacterial Cell Interactions with Nanopatterned Cicada Wing Surfaces. Biophysical Journal 104(4): 835–840. https://doi.org/10.1016/j.bpj.2012.12.046
And by “we all” I mean we, the eukaryotes, the organisms with complex cells with a nucleus, mitochondria and stuff.
The way organisms are classified changed hugely across the last two centuries but, during the past few decades, it became clear that we have three domains of life, namely Bacteria, Archaea and Eukarya. Although the relationship between these three domains was problematic at first, it soon became clear that Eukarya and Archaea are more closely related to each other than they are to Bacteria.
Both Bacteria and Archaea are characterized by the so-called prokaryotic cell, in which there is no delimited nucleus and only a single circular chromosome (plus a lot of smaller gene rings called plasmids). Eukarya, on the other hand, has a nucleus surrounded by a membrane which includes many linear chromosomes. Both the structure of the cell membrane and several genes indicate that Archaea and Eukarya are closely related, but it was still a mystery whether both groups evolved from a common ancestor and were, therefore, sister-groups, or whether eukaryotes evolved directly from archaeans and were, therefore, highy complex archaeans.
Things started to point toward the second hypothesis after several proteins originally considered exclusive to eukaryotes (the so-called Eukaryotic Signature Proteins, ESPs) were found in representatives of the clade TACK of Archaea. However, different clades within the TACK clade had different ESPs, so things remained uncertain.
Then in 2015 a new group of archaeans was discovered in the Arctic Ocean between Norway an Greenland near a field of active hydrothermal vents named Loki’s Castle (Spang et al. 2015). Named Lokiarchaeoata, this new archaean group contained a larger number of ESPs, including many found in different TACK lineages. Lokiarchaeota appeared as a sister-group of eukaryotes in phylogenetic reconstructions and indicated that eukaryotes evolved, indeed, from archaeans, and apparently from more complex archaeans than the ones known at the time. This group was solely based on an incomplete genome found in the sediments, as the organism itself was not found and could not be cultivated to confirm the structure of its cell.
In 2016, another new archaean lineage was discovered through a genome found in the White Oak River estuary on the Atlantic coast of the USA (Seitz et al., 2016). Named Thorarchaeota, this clade revealed to be closely related to Lokiarchaeota and, therefore, to Eukaryotes.
Reconstruction of possible metabolic routes found in Thorarchaeota based on the genes (white boxes) found in the thorarchaeotan genome. Credits to Seitz et al. (2016).
Then in 2017 a lot of new genomes were found in the same environments in which Lokiarchaeaota and Thorarchaeota had been found and in many others (Zaremba-Niedzwiedzka et al., 2017). They included two new groups closely related to these two, which were named Odinarchaeota and Heimdallarchaeota. This whole group received the name “Asgard archaeans” and phylogenetic reconstructions put Eukarya within it, with Heimdallarchaeota being Eukarya’s sister group.
But questions and doubts soon arised. Still in 2017, a new paper (Da Cunha et al., 2017) questioned these findings and raised the hypothesis that the phylogenetic reconstructions putting Asgard and Eukarya together was an artifact caused by long branch attraction, a side-effect of phylogenetic reconstructions in which fast-evolving species force distantly related clades to collapse into a single clade. The removal of some fast-evolving archaeans from the analysis was enough to break the Asgard-Eukarya relationship apart. Since the genomes of Lokiarchaeota and other Asgards were reconstructed from environmental DNA and not from single cells, there was a possibility that the samples were contaminated with material from other organisms. The protein genes used in the analyses also seemed to have divergent origins and may have been acquired via horizontal gene transfer, when a gene is transferred from one organism to another by means other than reproduction, usually through viruses.
The original authors of the Asgard clade, who proposed its proximity to Eukarya, rejected Da Cunha et al.’s (2017) criticism and stated that they used inadequate methodology and that there was no evidence of contamination in their samples (Spang et al., 2018).
(OMG, this turned into an actual fight. Grab your popcorns!)
Da Cunha et al. (2018) responded again showing more evidence of contamination and saying that Spang et al. should show evidence of inadequate methodology if it was the case.
Later studies continued to find the eukaryote sequences in new samples of Asgard, which decrease the likelihood of contamination (Narrowe et al., 2018).
Fournier & Poole (2018) discussed the implications of the proximity of Eukarya to Asgard and proposed a classification in which Asgard was not a member of Archaea anymore, but formed a new domain, Eukaryomorpha, together with Eukarya. They made an analogy with the mammals evolving from synapsids and how synapsids used to be seen as reptiles, even though they are not nested inside the Reptilia (Sauropsida) clade. The same would be the case of Asgard. Despite being “Archaea-like”, they would not be true archaeans.
A hypothetical topology of “true archaeans”, Asgard and Eukarya according to Fournier & Poole (2018).
In a study published in December, Williams et al. (2019) reanalyzed the issue using more data and recovered again the proximity of Asgard to Eukarya. With this accumulation of evidence, the hypothesis of Eukarya originating from inside Archaea grew stronger.
Then now, a few days ago, we finally got what we were waiting for. A group of Japanese scientists (Imachi et al., 2020) finally isolated an Asgard organism and was able to culture it in the lab. It was a very hard task, though. The culture grew very slowly, with a lag phase (the phase in which cells adapt to the environment and grow without dividing) lasting up to 60 days!
The creatures were growing in a mixed culture with a bacterium of the genus Halodesulfovibrio and an archaeon of the genus Methanogenium. The Asgard cells were named Candidatus Prometheoarcheum syntrophicum. In prokaryote taxonomy, a new species receives the status of Candidatus when it was not possible to maintain it in a stable culture.
The cells of this Asgard species are coccoid, i.e., spherical, and often present vesicles on the surface or long membrane protrusions that may or not branch. These protrustions do not connect to each other nor to other cells, differently from similar structures in other archaeans. The cells do not seem to contain any organelle-like structures inside them, going against the expectations. Asgard is not yet the eukaryote-like cell we were waiting for!
Several electron microscope images of Canidatus Prometheoarcheum syntrophicum. Vesicles show in e, f and proturision in g, h. Credits to Imachi et al. (2020).
Thanks to the culture of this Asgard species, it was possible to extract its whole genome and confirm what was previously known from Asgard and based solely on environmental DNA. This confirmed the presence of 80 ESPs and, in a phylogenetic analysis, this new species appeared as the sister group of Eukarya.
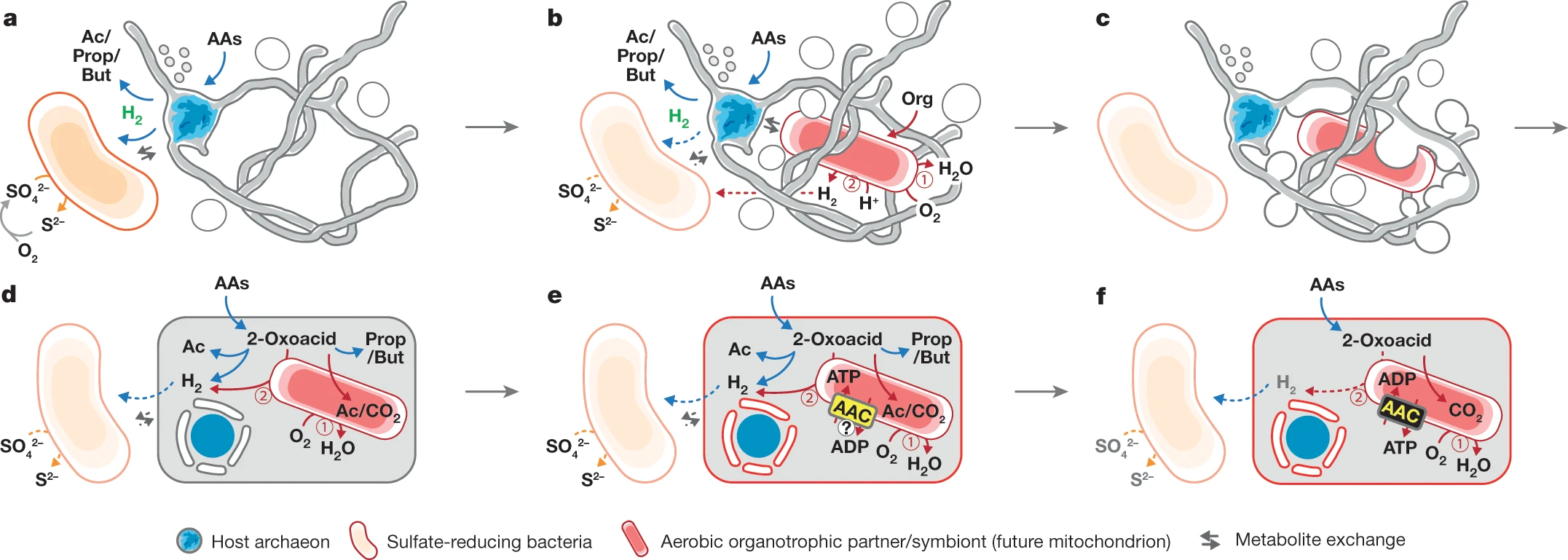
Candidatus Prometheoarcheum syntrophicumrevealed to be anaerobic and to feed on aminoacids, breaking them into fatty acids and hydrogen. Its association with the other two prokariotes in the mixed culture seems to be a sort of mutualism, with the three species helping each other by hydrogen transfer from one species to another. Many questions about how an organism like that turned into the complex eukaryotic cell still remain but at least we have some more hints about the acquisition of the mitochondria.
Hypothesis of eukaryotic cell evolution based on a mutualistic relationship between an Asgard-like archaean and an aerobic bacterium. Credits to Imachi et al. (2020).
The most widely accepted hypothesis was that primitive eukaryotic cells engulfed an aerobic bacteria through phagocytosis to eat it but ended up retaining it inside. However, seeing the cooperation of our Asgard archaean with other prokaryotes raises the hypothesis that maybe the mutualism between the pro-eukaryotic cell and the aerobic bacteria started when they were still separate organisms.
Are we ever going to find the “true” proto-eukaryote? Let’s wait for the next episodes.
Da Cunha V, Gaia M, Gadelle D, Nasir A, Forterre P (2017) Lokiarchaea are close relatives of Euryarchaeota, not bridging the gap between prokaryotes and eukaryotes. PLOS Genetics 13(6): e1006810. doi: 10.1371/journal.pgen.1006810
Da Cunha V, Gaia M, Nasir A, Forterre P (2018) Asgard archaea do not close the debate about the universal tree of life topology. PLOS Genetics 14(3): e1007215. doi: 10.1371/journal.pgen.1007215
Imachi H, Nobu MK, Nakahara N et al. (2020) Isolation of an archaeon at the prokaryote–eukaryote interface. Nature. doi: 10.1038/s41586-019-1916-6
Narrowe AB, Spang A, Stairs CW, Caceres EF, Baker BJ, Miller SC, Ettema TJG (2018) Complex Evolutionary History of Translation Elongation Factor 2 and Diphthamide Biosynthesis in Archaea and Parabasalids. Genome Biology and Evolution 10: 2380–2393. doi: 10.1093/gbe/evy154
Seitz KW, Lazar CS, Hinrichs KU, Teske AP, Baker BJ (2016) Genomic reconstruction of a novel, deeply branched sediment archaeal plylum with pathways for acetogenesis and sulfur reduction. ISME Journal 10: 1696–1705. doi: 10.1038/ismej.2015.233
Spang A, Saw JH, Jørgensen SL, et al. (2015) Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521: 173–179. doi: 10.1038/nature14447
Spang A, Eme L, Saw JH, Caceres EF, Zaremba-Niedzwiedzka K, et al. (2018) Asgard archaea are the closest prokaryotic relatives of eukaryotes. PLOS Genetics 14(3): e1007080. doi: 10.1371/journal.pgen.1007080
Williams TA, Cox CJ, Foster PG, Szőllősi GJ, Embley TM (2019) Phylogenomics provides robust support for a two-domains tree of life. Nature Ecology & Evolution. doi: 10.1038/s41559-019-1040-x
Zaremba-Niedzwiedzka K, Caceres EF, Saw JH et al. (2017) Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541:353–358. doi: 10.1038/nature21031
All around the world, many animal species have adapted to live in cave environments, places that are naturally devoid of light, either partially or entirely, and are, therefore, nutrient-poor habitats. The lack of light makes it impossible for plants and other photosynthetic organisms to survive and, as a result, little food is available for non-photosynthetic creatures. They rely almost entirely on food that enters the cave from the surface by water or animals that move between the surface and the depths.
Due to the lack of light in caves, animals adapted to this environment are usually eyeless, because seeing is not possible anyway, and white, because there is no need for pigmentation on the skin to protect from radiation or to inform anything visually. On the other hand, chemical senses such as smell and taste are often very well developed.
All these limitations make cave environments relatively species-poor when compared to surface environments. Or at least that is what it looks like at first. There are, of course, much less macroscopic species, such as multicellular animals, but those animals are themselves an environment and they may harbor a vast and unknown diversity of microrganisms inside them.
As you may know, most, if not all, animals have mutualistic relationships with microorganisms, especially bacteria, living in their guts. Those microorganisms are essential for many digestive processes and many nutrients that animals get from their food can only be obtained with the aid of those microscopic friends. The types of microorganisms in an animal’s gut are directly related to the animal’s diet. For example, herbivores usually have a high diversity of microorganisms that are able to break down carbohydrates, especially complex ones such as cellulose.
A recent study, conducted in China with fishes of the genus Sinocyclocheilus, compared the gut microbial diversity of different species, including some that live on the surface and some that are adapted to caves. All species of Sinocyclocheilus seem to be primarily omnivores but different species may have preferences for a particular type of food, being more carnivorous or more herbivorous.
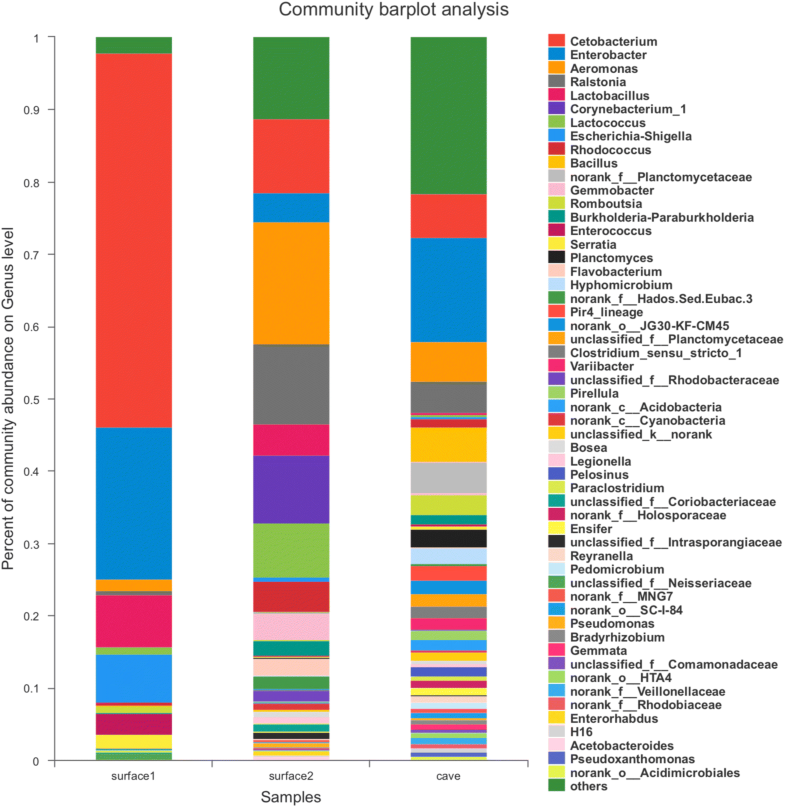
The study found that cave species of Sinocyclocheilus have a much higher microbial diversity than surface species. But how can this be possible if there is a limited number of resources available in caves compared to the surface? Well, that seems to be exactly the reason.
Sinocyclocheilus microphthalmus, one of the cave-dwelling species used in this study. Photo extracted from the Cool Goby Blog.
As I mentioned, species of Sinocyclocheilus are omnivores. On the surface, they have plenty of food available and can have the luxury of choosing a preferred food type. As a result, their gut microbiome is composed mainly by species that aid in the digestions of that specific type of food. In caves, on the other hand, food is so scarce that one cannot chose and must eat whatever is available. This includes feeding on small amounts of many different food types, including other animals that live in the cave and many different types of animal and plant debris that reach the cave through the water. Thus, a much more diverse community of gut microorganisms is necessary for digestion to be efficient.
Look how the number of different genera of bacteria is much larger in the cave group (right) than in two groups of surface species (left and center). Image extracted from Chen et al. (2019).
More than only an increased diversity by itself, the gut community of cave fish also showed a larger number of bacteria that are able to neutralize toxic compounds of several types. The reason for this is not clear yet but there are two possible explanations that are not necessarily mutually exclusive. The first states that water in caves is renewed in a much lower rate than surface waters, which promotes the accumulation of all sort of substances, including metabolic residues of the cave species themselves that can be toxic. The second hypothesis is of greater concern and suggests that this increased number of bacteria that are able to degrade harmful substances is a recent phenomenon caused by an increase in water pollutants coming from human activities, which is promoting a selective pressure on cave organisms.
The diverse gut microbiome of cave fish is, therefore, a desperate but clever strategy to survive in such a harsh environment. Nature always finds a way.
Chen H, Li C, Liu T, Chen S, Xiao H (2019) A Metagenomic Study of Intestinal Microbial Diversity in Relation to Feeding Habits of Surface and Cave-Dwelling Sinocyclocheilus Species. Microbial Ecology. doi: 10.1007/s00248-019-01409-4
With the current raise in genetically modified crops and all controversies around them, you probably heard about things such as Bt corn and Bt cotton. But do you know what Bt means?
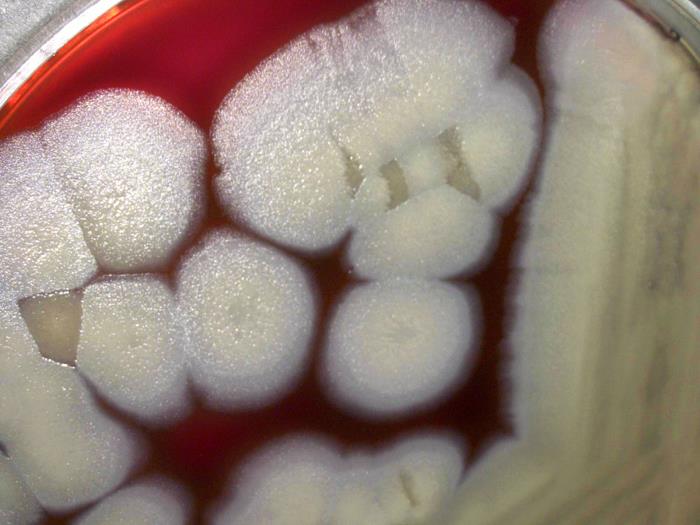
Bt stands for Bacillus thuringiensis, and this is going to be our fellow for today. A Gram-positive bacterium, Bt is found in many environments, including the soil, the surface of several plants and in the gut of several species of caterpillars. Belonging to the large and heterogeneous genus Bacillus, Bt is very closely related to, and sometimes considered as being of the same as, Bacillus cereus, which can can foodborne illness, and Bacillus anthracis, the species that causes anthrax. The main differences between these three species relies on their plasmids (small DNA pieces in a bacterium’s cytoplasm), while the genetic composition of their chromosome is basically the same.
Colonies of Bacillus thuringiensis growing on sheep blood agar.
As all species of Bacillus, Bt can sporulate, i.e., convert itself into a dormant form called endospore (sometimes wrongly named spore) when environmental conditions are not favorable. During sporulation, Bt forms crystals of delta-endotoxins, a proteinaceous inseticide also named crystal proteins or cry proteins. Cry proteins are encoded by cry genes, which are located in plasmids and not in the bacterial chromosome. When insects and nematodes ingest those crystals, they are denatured in the alkaline environment of the animal’s gut and become soluble. In soluble phase, the crystals are digested and liberate their toxins, which then paralyze the digestive tract and make the animal starve to death. The number of different cry proteins in Bt is really large, indicating a yet unknown selective pressure to the development of such a sophisticated defense mechanism.
Gram-stained colony of Bt under a 1000 X magnification. Photo by Wikimedia user Dr. Sahay.*
Due to this strong insecticidal effect, Bt endospores and cry proteins have been used to control insect pests during the last century. The insecticide is usually applied as a spray and can be bought under different trademarks. However, because of natural selection, the pests end up developing resistance to the toxins and new strains of Bt are constantly produced to originate newer varieties of the insecticide.
Since the 1980s, there have been studies on the production of genetically modified organisms that incorporate Bt genes responsible for the production of cry proteins. Currently, the two most widely cultivated genetically modified crops containing Bt genes are Bt corn and Bt cotton. The cry genes of Bt have been introduced in the DNA of these species, allowing them to synthesize cry proteins. When an insect eats the leaves of such plants it ingests the crystals and dies as if it would have eaten the spores of Bt. And in fact, that is what happened. The introduction of Bt corn, for example, strongly reduced the attack of several corn pests, such as the corn borer.
Although the use of Genetically Modified Organisms (GMOs) is still seen as bad by many people, most studies have shown that they are relatively safe compared to many other human-interference activities. Bt corn and Bt cotton were shown to be safe for non-targeted organisms and to the environment as a whole. The main problem with GMOs is the fact that the technology to produce them lies in the hands of giant profit-maker companies.
Helgason, E.; Økstand, O. A.; Caugant, D. A.; Johansen, H. A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.-B. (2000) Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis — One species on the basis of genetic evidence. Applied and Environmental Microbiology, 66(6): 2627–2630. doi: 10.1128/AEM.66.6.2627-2630.2000
Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D. R.; Dean, D. H. (1998) Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Reviews, 62(3): 775–806.
The success of many plant species wordlwide is the result of their association with different organisms in their root systems, such as fungi and bacteria. Among bacteria, the most widely known group of root-associated species is that of the so-called rhizobia, bacteria that are associated with the root of legumes (plants of the family Fabaceae).
However, this kind of association evolved independently many times in many lineages of plants and many lineages of bacteria. Today I’ll present you a bacterium that is not closely related to the rhizobia, but acts similarly. Known scientifically as Frankia alni, it does not have a common name, but I decided to call it the alder-root bacterium.
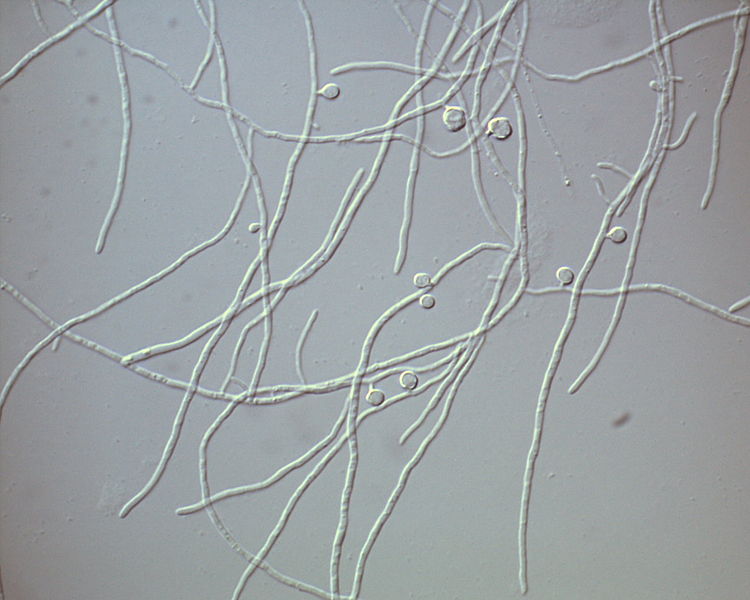
As both its scientific name and its brand-new common name imply, the alder-root bacterium is a species associated with the roots of trees in the genus Alnus, commonly known as alder in English. It belongs to the phylum Actinobacteria and the order Actinomycetales, having a filamentous growth that produces a structure similar to the mycelia of fungi.
The hyphae that form the mycelium of Frankia alni. Photo by Wikimedia user Kkucho.*
The bacteria do not penetrate the cell membranes of the host plant at first, but remain inside a structure that develops from the plant cell wall. From there, they stimulate cell division and the production of nodules that grow from the rooth and inside of which they migrate, entering the nodule’s cells.
A nodule caused by Frankia alni on the root of an alder species. Photo by Gerhard Schuster.*
Through a complex biochemical process that I won’t present here in detail, the alder-root bacteria can capture nitrogen from the atmosphere and synthesize aminoacids from it, part of which is shared with the host. As a result, alder trees that contain nodules of alder-root bacteria are able to grow in nitrogen-poor soils and eventually enrich that soil and allow other plants to establish.
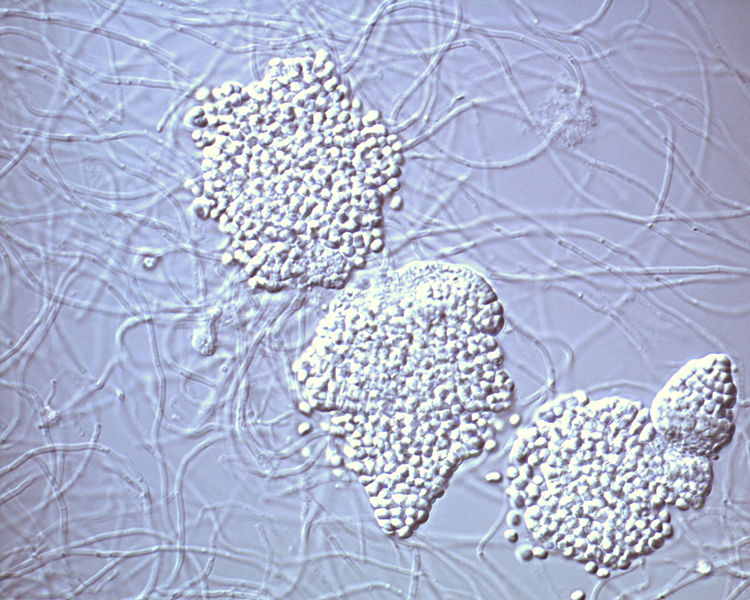
Sporangia of Frankia alni. Photo by Wikimedia user Kkucho.*
In order to disperse themselves through the environment and find new hosts, the alder-root bacteria produce spores inside sporangia. Once released, the spores may migrate, probably through water, to new localities where they germinate and restart the cycle.
Today we’ll return to the tiny world of the bacteria once more. And I guess it is a good time to introduce another celebrity from the bacterial world, the hay bacillus or grass bacillus, Bacillus subtilis.
Several colonies of Bacillus subtilis on agar. Photo by Wikimedia user Debivort.*
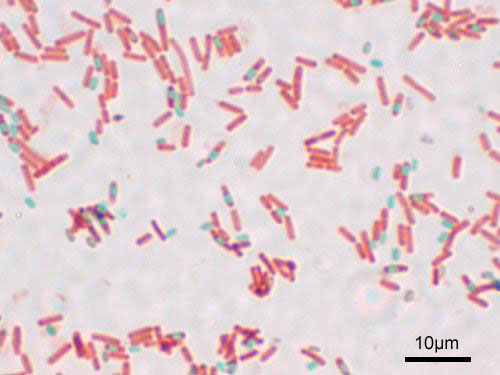
As any typical Bacillus, the hay bacillus has rod-shaped cells, hence the name. They measure about 4–10 µm in length and 0.25–1.0 µm in diameter and have many flagella, so they can move quickly in a liquid medium. The natural habitat of the hay bacillus is the soil, but it can also be found in the intestine of mammals, including humans.
As it is common among the members of the phylum Firmicutes, the hay bacillus is able to enter in a dormant form called endospore that is able to tolerate extreme environmental conditions. They can survive in this form for decades, centuries, perhaps even millenia, until the conditions are adequate again.
Microscopic image showing vegetative (red) and endospores (green) of Bacillus subtilis. Photo by Wikimedia user Y tambe.*
The hay bacillus is one of the most studied and cultivated bacterium in the world, being considered a model organism. In East Asia, one of its varieties is used in the production of the Japanese traditional food nattō. Before the introduction of antibiotics, it was common to use cultures of B. subtilis in treatments to improve immunological responses. Currently, it is used in laboratory studies focused on the formation of endospores and the phenomenon of transformation, a process by which a bacterium can capture DNA from the medium in which it is and incorporate it into its own genetic material. Additionally, it is used to produce a variety of substances, including naturally produced antibiotics.




 This work is licensed under a Creative Commons Attribution 4.0 International License.
This work is licensed under a Creative Commons Attribution 4.0 International License.
 This work is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.
This work is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.